 2015, Vol. 34
2015, Vol. 34文章信息
- 周毅, 郑猛猛, 卢松贺, 程昕昕, 蒋德, 汪建飞, 刘正
- ZHOU Yi, ZHENG Meng-meng, LU Song-he, CHENG Xin-xin, JIANG De, WANG Jian-fei, LIU Zheng
- 硫酸锌处理对玉米幼苗养分和锌累积的影响
- Effects of Zinc Sulfate on Nitrogen, Phosphorus, Potassium and Zinc Accumulation in Different Parts of Maize(Zea mays L.) Seedlings at Germination Stage
- 农业环境科学学报, 2015, 34(1): 29-36
- Journal of Agro-Environment Science, 2015, 34(1): 29-36
- http://dx.doi.org/10.11654/jaes.2015.01.005
-
文章历史
- 收稿日期:2014-05-06

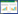



2. 玉米育种安徽省工程技术研究中心, 安徽 凤阳 233100;
3. 华南理工大学环境与能源学院, 广州 510641
2. Institute of Anhui Engineering Technology on Maize Breeding, Fengyang 233100, China;
3. College of Environment and Energy, South China University of Technology, Guangzhou 510641, China
锌是兼具营养和毒性的动植物必需元素之一,也是唯一的在6大酶类结构组成中均存在的金属元素[1],缺锌或锌过量均可导致动植物出现不良症状[1, 2, 3, 4]。我国土壤的锌含量为3~790 mg·kg-1,平均含量为100 mg·kg-1 [6]。由于土壤锌含量低于200 mg·kg-1时,对植物生长[5]、土壤微生物数量及酶活性[7]无不良效应,我国土壤理应很少出现锌过量的问题。近年来,伴随锌在钢铁防腐、合金制造、航空航天器保护层以及电子系统的电路保护等领域的广泛应用[8],在锌矿周边地区及流域,已陆续出现复杂的环境问题[2, 4, 7, 8, 9],严重时甚至导致水生生物死亡[2]、农作物绝收和地块荒芜[8]。另外,持续施用含有锌副产物的磷肥或有机肥料等也可能存在影响土壤和农作物产品质量的风险[10]。目前,我国并没有针对商品有机肥[10]和食品锌含量的相关限量标准,但已有研究证实,硫酸锌的浸种浓度低于800 mg·L-1时能显著提高玉米种子的萌发速率,当浓度升高至1600 mg·L-1时其促进效应开始下降[11],说明该浓度接近引起玉米种子萌发的抑制效应的临界浓度。本文拟通过以低于1600 mg·L-1的不同浓度硫酸锌溶液浸种模拟临界锌过量条件,研究硫酸锌对萌芽阶段玉米胚芽、胚根“作物三要素”(氮、磷、和钾)累积量以及胚乳氮、磷和钾残余量动态变化的影响及其与锌累积的关系,为揭示锌污染生物毒性效应的营养生理机制,实现含锌类肥料的安全应用,以及为临界锌污染生物测试评价体系的建立提供参考。 1 材料与方法 1.1 实验方法
采用单因素5水平实验设计。以ZnSO4·7H2O溶液模拟锌污染。ZnSO4·7H2O浓度分别为0(蒸馏水,对照)、400、800、1200、1600 mg·L-1,每个水平(处理)设16个重复。玉米种子(郑单958)放置在垫有滤纸的方形发芽盒(长×宽×高=12 cm×12 cm×5 cm)中。每个发芽盒选取100粒大小均匀、籽粒饱满的玉米种子,加50 mL ZnSO4溶液。称量每个发芽盒的初始重量并记录。将发芽盒置于25 ℃培养箱中进行暗培养。当萌芽明显,胚芽尖呈现黄色时,揭盖进行光照培养(光暗比10 h∶14 h)。分别在第1、4、7、10 d采样,每次采样随机取4个重复,并对剩余发芽盒分别补充蒸馏水至初始重量。 1.2 测定项目与方法 1.2.1 玉米幼苗胚根、胚芽生物量干重和胚乳干物质量测定
分别洗涤不同处理的玉米幼苗,擦干表面水分,随机选取20株,称重后将每个培养皿中的幼苗分胚根、胚芽和胚乳3部分。于105~110 ℃杀青30 min后,再于70~80 ℃的烘箱中烘至恒质量,分别得到胚根、胚芽的生物量干重(生物量累积量)和胚乳干物质量。 1.2.2 植物样品全氮、磷、钾和锌含量的测定
植物烘干样品粉碎后采用H2SO4-H2O2法消煮,溶液中的全氮含量采用流动分析仪(AA3,美国BRAN+LUEBBE公司)测定;全磷含量采用磷钼蓝比色法测定;全钾含量采用火焰光度计法测定;另取适量植物样品经干灰化法灰化后,采用硝酸溶解灰分,溶液中的全锌含量直接用ICP(7000DV,美国PerkinElmer 公司)法测定。氮(磷、钾或锌)累积量=全氮(磷、钾或锌)含量×生物量干重;氮(磷、钾或锌)残余量=全氮(磷、钾或锌)含量×生物量干重。 1.3 数据处理
试验数据采用Excel 2003和SPSS 19.0统计软件进行制图、变量间线性回归方程的配置与相关分析。 2 结果与分析 2.1 硫酸锌对萌芽阶段玉米胚根、胚芽生物量和胚乳干物质残余量的影响
如图 1所示,在不同锌浓度下,随着发芽天数的延长,玉米胚根和胚芽的生物量均呈近似线性增长的趋势,而其胚乳干物质量则呈近似线性降低的趋势。分别配置不同锌浓度下,发芽天数与胚根、胚芽生物量和胚乳干物质量的线性回归方程。如表 1所示,当锌浓度在400 mg·L-1或800 mg·L-1时,不同部位回归方程的斜率的绝对值出现峰值,说明低浓度锌可能具有刺激玉米胚乳贮藏物质转化并运输至胚芽和胚根、加速其生长的效应。当锌浓度大于800 mg·L-1浓度时,则出现胚芽和胚根生物量累积量低于对照,而胚乳的干物质残余量高于对照的现象,且其回归方程的斜率的绝对值也均出现低于对照的现象,说明高浓度硫酸锌可能抑制胚乳贮藏物质转化及向胚芽和胚根的运输,从而抑制其生长。

|
| 图 1 不同Cd含量处理下,菌剂对水稻株高的影响 Figure 1 Effect of AMF on rice plant height under different concentrations of Cd |
 |
锌浓度对玉米幼苗氮、磷和钾含量的影响不明显。由图 2可以看出,在不同锌浓度下,胚根(钾累积量除外)与胚芽氮、磷和钾的累积量,及胚乳中这3个元素的残余量的变化规律均与其生物量累积量、残余量的变化规律基本一致,即随着发芽天数的延长,呈近似线性增长或近似线性降低的趋势,说明低浓度硫酸锌具有刺激胚芽和胚根累积养分的效应。当锌浓度大于800 mg·L-1浓度时,也出现胚芽和胚根养分累积量低于对照(蒸馏水培养)而胚乳养分残余量高于对照的现象,说明高浓度硫酸锌可能通过抑制胚乳中的养分转化及向胚芽和胚根的运输,抑制胚芽和胚根的生长。

|
| 图 2 硫酸锌对萌芽阶段玉米胚芽、胚根养分含量、累积量和胚乳养分含量、残余量的影响 Figure 2 Effect of ZnSO4 on content and accumulation/residue of nutrients in germ,radicle and endosperm of maize at germination stage |
在萌发第10 d,3种养分中以胚乳钾残余量的降低幅度最大,其次为磷,说明种子中转化速率最快的是钾。但是,在不同锌浓度条件下,胚根钾累积量的线性回归方程F检验差异均不显著。由图 1可以看出,在第4~7 d,胚乳养分的耗竭速率均存在减缓的趋势,此时胚根养分累积量基本保持不变或略有降低,胚芽养分累积量的升高趋势也有所减缓,但其变化幅度不及胚根明显。第7 d后,胚根和胚芽的养分累积速率明显回升,而对应胚乳的氮和磷的消耗也开始有所加快,且在第10 d,随着锌浓度的增加,胚乳氮的残余量逐渐增多。因此,在第4~7 d,胚乳养分向胚芽和胚根的运输可能出现出现阻滞,且高浓度锌可能明显影响该阶段之后的胚乳氮的转化。目前还不清楚其内在的机制,尚有待进一步的研究。 2.3 硫酸锌对萌芽阶段玉米不同部位锌累积的影响
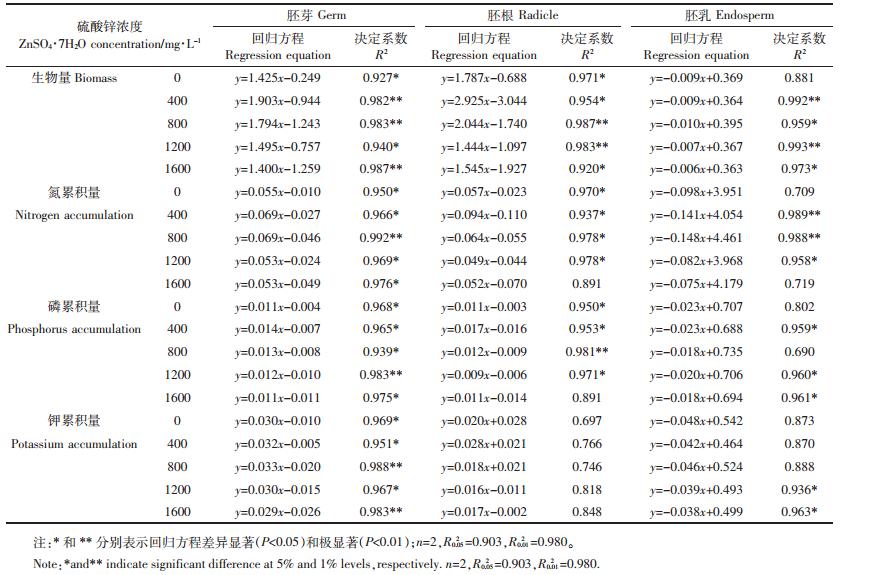
如图 3所示,在第10 d,随锌浓度的增加,胚芽、胚根和胚乳中的锌含量和累积量均呈线性升高的趋势,且其线性回归方程均达差异极显著水平(P<0.01),表明在高硫酸锌浓度下,胚芽和胚根的生物量、氮、磷和钾累积量低于对照,可能与其锌含量及累积量的持续升高有关。锌累积量回归方程的斜率由高到低依次为胚乳、胚根和胚芽,锌含量回归方程的斜率由高到低依次为胚根、胚芽和胚乳。

|
| 图 3 硫酸锌对萌芽阶段玉米胚芽、胚根和胚乳锌累积的影响 Figure 3 Effect of ZnSO4 on Zn accumulation in germ,radicle and endosperm of maize at germination stage |
如表 2所示,不同处理的胚乳锌累积占整株总锌累积的百分率明显高于胚根和胚芽,对照处理的胚芽锌累积百分率相对高于胚根,而在其他锌浓度下,胚根的锌累积百分率则高于胚芽。
 |
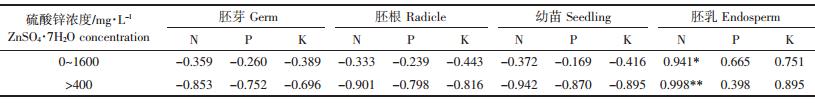
由表 3可见,虽然锌浓度高于400 mg·L-1时,氮、磷和钾累积与锌累积的相关系数明显大于试验设置的整体浓度范围(0~1600 mg·L-1)内的相关系数,但其中仍然只有胚乳氮的残余量与其锌累积量呈显著正相关关系。这说明在籽粒萌发过程中,胚乳锌累积量的增加可能通过抑制种子中的氮素转化降低了胚乳中的氮消耗速率,对此尚有待进一步研究证实。
 |
玉米是对锌敏感的作物之一[12]。已有研究表明,适量的锌处理可显著提高种子的萌发速率、萌发过程的抗氧化水平[11]和物质代谢水平,但超过一定剂量则出现抑制作用[11, 13]。一般认为,对高浓度锌毒害的最敏感部位是植物的根系[14, 15],因为锌向地上部的迁移相对较难,故而主要累积在植物根部并使其严重受害[16]。Mahmood等[13]和保琼莉等[6]则认为,植株地上部锌含量和吸收量均高于根系,其受抑制的程度也高于根系,因而呈现出随锌浓度的增加,根冠比明显升高的趋势[12],且当锌浓度进一步增大时,根冠比在出现峰值后下降所达的数值仍明显高于其对照[6]。
在本实验中,由能够表征生长速率的生物量累积量动态变化的回归方程的斜率可以看出,在相同的锌浓度下,胚根和胚芽的生长速率各不相同,说明在萌发阶段,胚根和胚芽可能也具有阶段发育的特性,但二者的发育并不同步。这可能是种子养分含量与转化速率等内因对胚根和胚芽生长的影响存在程度不等的交互效应的结果。因此,是否能以萌发阶段幼苗根冠比的变化评价胚根和胚芽对锌的促进或毒性效应的相对敏感性,尚有待商榷。本实验中除对照外,在不同锌浓度下胚根的锌含量均明显高于胚芽(结果未列)。与前人研究结果出现明显差异的原因可能是:本实验与上述研究采用的锌浓度、单个植株承受的锌的容量(锌浓度和溶液体积的乘积),以及处理设置开始及持续的时间不同。以锌浓度和ZnSO4·7H2O浓度的换算系数(为4.40)计算,Mahmood等[13](3~12 mg·L-1)和保琼莉等[6](0~40 mg·L-1)实验设置的锌浓度均明显低于本实验设置的浓度(0~364 mg·L-1);同时,也不能排除在Mahmood等和保琼莉等的研究中,存在地上部从茎、叶基部接触到的营养液(浸没种子的营养液或固定植株的棉花所吸附的营养液)直接吸收锌而导致地上部出现较高锌累积的可能性。这也可以从叶片喷施锌对提高籽粒锌含量的效应明显高于土施锌[17, 18]以及锌在韧皮部的再转移能力较弱[16]等研究结果得到证实,即茎或叶基部途径吸收的锌可能是造成地上部锌累积程度较高的主要原因之一。 3.2 锌污染对萌发阶段玉米氮、磷和钾营养的毒性效应特征
已有研究认为,重金属在低剂量作用时可促进植物过量生成活性氧[19, 20, 21, 22]和活性氮,进而通过轻微的加强氧化应激反应对植物的萌发、生长等产生一定的刺激作用[19]。在本实验中,当锌浓度在400 mg·L-1和800 mg·L-1时,胚芽和胚根的氮、磷和钾累积量以及胚乳的氮、磷和钾残余量的回归方程的斜率均高于对照,证实低浓度硫酸锌具有刺激玉米胚乳贮藏物质转化,加速胚芽和胚根生长及累积氮、磷和钾的效应。
Gomes等[20]认为,过高浓度的锌具有降低萌发率或延迟萌发的效应,因为重金属可能通过直接的毒性效应或抑制吸涨过程中的水分吸收两个途径对种子的萌发产生影响[15]。但Lefevre等[19]认为,锌不影响种子的吸涨过程,在高浓度锌溶液中吸涨的种子,即使转移至去离子水中,仍然难以萌发,因为活性氧对种子中贮存的蛋白质、脂类等物质所造成的氧化性损伤可能是重金属对种子萌发产生毒性效应的主要原因。已有研究证实,在细胞水平上,过量锌可改变有丝分裂活性[21],加速活性氧的生成[19],导致膜脂过氧化程度加重[22],不仅影响膜的完整性和透性[21],而且导致蛋白质发生氧化羰基化修饰,不可逆失活或发生交联,更易于被蛋白酶降解,呼吸电子传递链的NADH脱氢酶和氧化磷酸化受到抑制[22],淀粉水解酶和酸性磷酸酶活性下降[19]等一系列新陈代谢的过程发生改变,甚至出现细胞死亡[22],显著改变萌发过程中种子贮藏物质的转化及向胚根、胚芽中的运输。
在本实验中,当锌浓度大于800 mg·L-1时,胚芽和胚根生物量和养分的累积速率以及胚乳干物质的耗竭速率均开始下降,证实高浓度硫酸锌对胚乳、胚芽和胚根中的物质转化存在明显的抑制效应。尽管实验未同时测定蛋白质、脂肪和淀粉等物质的含量及其相应的水解酶活性变化,但在元素水平上,可由氮、磷和钾含量与累积量(或残余量)的变化及其与锌累积量变化的相关关系看出:在种子萌发过程中,胚乳锌累积量的增加可能通过抑制种子中的氮素转化减缓了胚乳中的氮消耗。目前,还不清楚萌发过程中胚乳中锌累积的发生及其动态变化的内在机制,相关的研究也少见报道,有待进一步的研究。 4 结论
低浓度硫酸锌具有刺激玉米胚乳、胚芽和胚根中物质转化,促进幼苗氮、磷和钾累积和植株生长的效应。胚芽、胚根和胚乳中的锌含量及锌累积量随硫酸锌浓度的增加均呈线性增高的趋势。当硫酸锌浓度超过800 mg·L-1时,对胚乳、胚芽和胚根中的物质转化存在明显的抑制效应。不同部位的锌累积速率及其锌累积占整株锌总累积量的百分率由高到低的次序均为胚乳、胚根和胚芽,胚乳的高锌累积导致萌发后期种子氮的消耗速率减慢。种子贮存的3种养分中,钾的转化速率最快。在种子萌发的第4~7 d,不同硫酸锌浓度条件下胚乳养分的消耗速率均呈减慢的趋势,对应胚根和胚芽的养分累积速率基本保持不变或略有降低,且此时胚芽氮、磷和钾的累积速率减小的幅度不及胚根明显,进一步证实胚根对锌污染相对较敏感。
| [1] | Long X X, Chen X M, Wong J W C, et al. Feasibility of enhanced phytoextraction of Zn contaminated soil with Zn mobilizing and plant growth promoting endophytic bacteria[J]. Trans actions of Nonferrous Metals Society of China, 2013, 23:2389-2396. |
| [2] | 李丽娜, 陈振楼, 张亚雷, 等. 长江口滨岸潮滩底栖动物泥螺受锌污染的急性毒理试验研究[J]. 生命科学研究, 2008, 12(4):373-376. LI Li-na, CHEN Zhen-lou, ZHANG Ya-lei, et al. Acute toxicity test of benthic Fauna Bullacta Exarata by pollutant zinc in the tidal flat of Changjiang estuary[J]. Life Science Research, 2008, 12(4):373-376. |
| [3] | 林 蕾, 陈世宝. 土壤中锌的形态转化、影响因素及其有效性研究进展[J]. 农业环境科学学报, 2012, 31(2):221-229. LIN Lei, CHEN Shi-bao. Transformation and influence factors of speciation of zinc in soils and its effect on zinc bioavailability:A review[J]. Journal of Agro-Environment Science, 2012, 31(2):221-229. |
| [4] | 罗旺兴, 叶挺进, 黄禹坤, 等. PPC 去除水源水中突发性重金属铜和锌污染研究[J]. 供水技术, 2012, 6(5):22-26. LUO Wang-xing, YE Ting-jin, HUANG Yu-kun, et al. Removal of emergent contamination of Cu2+ and Zn2+ in source water by potassium permanganate composites[J]. Water Technology, 2012, 6(5):22-26. |
| [5] | 刘 铮. 我国土壤中锌含量的分布规律[J]. 中国农业科学, 1994, 27(1):30-37. LIU Zheng. Regularities of content and distribution of zinc in soils of China[J]. Scientia Agricultura Sinica, 1994, 27(1):30-37. |
| [6] | 保琼莉, 田霄鸿, 杨习文, 等. 不同供Zn量对三种小麦基因型幼苗生长和养分吸收的影响[J]. 植物营养与肥料学报, 2007, 13(5):816-823. BAO Qiong-li, TIAN Xiao-hong, YANG Xi-wen, et al. Effects of different Zn supply rates on growth and nutrient uptake of wheat seedlings of three genotypes under the condition of solution culture[J]. Plant Nutrition and Fertilizer Science, 2007, 13(5):816-823. |
| [7] | 李 伟, 韦晶晶, 刘爱民, 等. 吊兰生长对锌污染土壤微生物数量及土壤酶活性的影响[J]. 水土保持学报, 2013, 27(2):276 -281. LI Wei, WEI Jing-jing, LIU Ai-min, et al. Effects of chlorophytum comosum growth on microbial biomass and soil enzymatic activities in zinc-polluted soil[J]. Journal of Soil and Water Conservation, 2013, 27(2):276-281. |
| [8] | 李 雷, 姜 郁, 夏新媛, 等. 我国锌工业可持续发展策略研究[J]. 有色金属工程, 2011(2):43-45. LI Lei, JIANG Yu, XIA Xin-yuan, et al. Research on the sustainable development of Chinese zinc industry[J]. Nonferrous Metal Engineering, 2011(2):43-45. |
| [9] | 王晓华, 邓喀中, 杨化超, 等. 土壤重金属污染信息提取遥感模型的建立:以水口山矿区铅锌污染为例[J]. 测绘通报, 2013(3):29-31. WANG Xiao-hua, DENG Ka-zhong, YANG Hua-chao, et al. Building-up of remote sensing models for heavy metal pollution in soil:Take the pollution of lead and zinc mine in Shuikou mountain as an example[J]. Bulletin of Surveying and Mapping, 2013(3):29-31. |
| [10] | 王 美, 李书田. 肥料重金属含量状况及施肥对土壤和作物重金属富集的影响[J]. 植物营养与肥料学报, 2014, 20(2):466-480. WANG Mei, LI Shu-tian. Heavy metals in fertilizers and effect of the fertilization on heavy metal accumulation in soils and crops[J]. Journal of Plant Nutrition and Fertilizer, 2014, 20(2):466-480. |
| [11] | 杨建肖, 杨国航, 孙世贤, 等. 硫酸锌处理对玉米种子萌发的生理效应[J]. 植物营养与肥料学报, 2009, 15(2):410-415. YANG Jian-xiao, YANG Guo-hang, SUN Shi-xian, et al. Physiological effects of zinc sulphate on maize seed germination[J]. Plant Nutrition and Fertilizer Science, 2009, 15(2):410-415. |
| [12] | 严小龙, 张福锁. 植物营养遗传学[M]. 北京:中国农业出版社, 1997:153-54. YAN Xiao-long, ZHANG Fu-suo. Plant nutrition genetics[M]. Beijing:Chinese Agriculture Press, 1997:153-154. |
| [13] | Mahmood S, Hussain A, Saeed Z. et al. Germination and seedling growth of corn(Zea mays L.) under varying levels of copper and zinc[J]. International Journal of Environmental Sciences and Technology, 2005, 2(3):269-274. |
| [14] | Marichalia A, Dallalia S, Ouerghemmia S, et al. Germination, morpho-physiological and biochemical responses of coriander(Coriandrum sativum L.) to zinc excess[J]. Industrial Crops and Products, 2014, 55:248-257. |
| [15] | Kranner I, Colville L. Metals and seeds:Biochemical and molecular implications and their significance for seed germination[J]. Environmental and Experimental Botany, 2011, 72(1):93-105. |
| [16] | 汪 洪, 金继运. 植物对锌吸收运输及积累的生理与分子机制[J]. 植物营养与肥料学报, 2009, 15(1):225 -235. WANG Hong, JIN Ji-yun. The physiological and molecular of zinc uptake, transport, and hyper-accumulation in plants:A review[J]. Plant Nutrition and Fertilizer Science, 2009, 15(1):225 -235. |
| [17] | 曹玉贤, 田霄鸿, 杨习文, 等. 土施和喷施锌肥对冬小麦籽粒锌含量及生物有效性的影响[J]. 植物营养与肥料学报, 2010, 16(6):1394-1401. CAO Yu-xian, TIAN Xiao-hong, YANG Xi-wen, et al. Effects of soil and foliar applications of Zn on winter wheat grain Zn concentration and bioavailability[J]. Plant Nutrition and Fertilizer Science, 2010, 16(6):1394 -1401. |
| [18] | 齐义涛, 周三妮, 张 庆, 等. 结实期叶面施锌对小麦籽粒不同部位锌生物有效性的影响[J]. 农业环境科学学报, 2013, 32(6):1085-1091. QI Yi-tao, ZHOU San-ni, ZHANG Qing, et al. The effect of foliar Zn application at grain filling stage on Zn bioavailability in grain fractions of modern winter wheat cultivars[J]. Journal of Agro-Environment Science, 2013, 32(6):1085-1091. |
| [19] | Lefevre I, Marchal G, Correal E, et al. Variation in response to heavy metals during vegetative growth in Dorycnium pentaphyllum Scop[J]. Plant Growth Regulation, 2009, 59(1):1-11. |
| [20] | Gomes M P, Carneiro M M L C, Nogueira C O G, et al. The system modulating ROS content in germinating seeds of two Brazilian savanna tree species exposed to As and Zn[J]. Acta Physiologiae Plantarum, 2013, 35(4):1011-1022. |
| [21] | Pavlíková D, Pavlík M, Procházková D, et al. Nitrogen metabolism and gas exchange parameters associated with zinc stress in tobacco expressing an ipt gene for cytokinin synthesis[J]. Journal of Plant Physiology, 2014, 171(7):559-564. |
| [22] | Gutierrez-Carbonell E, Lattanzio G, Sagardoy R, et al. Changes induced by zinc toxicity in the 2-DE protein profile of sugar beet roots[J]. Journal of Proteomics, 2013, 94:149-161. |