 2016, Vol. 35
2016, Vol. 35文章信息
- 臧怀敏, 李刚, 修伟明, 魏琳琳, 倪土, 杨殿林, 赵建宁
- ZANG Huai-min, LI Gang, XIU Wei-ming, WEI Lin-lin, NI Tu, YANG Dian-lin, ZHAO Jian-ning
- 磷高效转基因水稻OsPT4种植对土壤细菌群落多样性的影响
- Diversity of bacterial community in rice paddy soil grown with P-efficient transgenic rice(OsPT4)
- 农业环境科学学报, 2016, 35(3): 500-506
- Journal of Agro-Environment Science, 2016, 35(3): 500-506
- http://dx.doi.org/10.11654/jaes.2016.03.013
-
文章历史
- 收稿日期: 2015-10-14





从1996年到2014年,全球转基因作物的种植面积增加了100倍以上,转基因作物已经成为现代农业史上推广最为迅速的农作物[1]。据统计,2014年全球有28个国家种植转基因作物,面积超过1.81亿hm2。但是转基因作物在给人们带来巨大经济效益的同时,其所带来的生态安全性问题也日益引起公众的广泛关注[2, 3, 4]。
磷高效转基因作物是通过生物技术手段培育出的磷高效吸收、转运的作物新品种,用以克服因土壤缺磷造成的作物缺磷问题[5],与常规作物相比,其磷素吸收、转运效率得到了显著提高[6]。磷高效转基因作物因特有的磷素高效吸收和转运能力可以使其在土体有效磷浓度较低的情况下吸收到更多的磷素,从而满足作物的正常生长发育需要。磷高效转基因水稻OsPT4为南京农业大学资源与环境学院植物营养分子生物学实验室通过转基因技术研制的磷高效吸收水稻新材料,是通过克隆水稻家族的pht1家族成员OsPT4基因,并将其导入日本晴而获得的超表达材料,在水培正常供磷条件下,地下部全磷含量较对照增加10%~20%,地上部全磷含量增加36%~50%;在水培低磷条件下,地下部全磷含量较对照增加8%~15%,地上部全磷含量增加15%~30%[7, 8]。磷高效突变体材料PHO2是编码OsLTN1基因发生无义突变产生的磷高效吸收材料,该突变体材料增加了水稻磷素的吸收和转运能力,并且使磷素在新叶部位累积[9]。
土壤磷库长期处在一个动态的平衡状态,磷高效转基因水稻对土壤磷素高效吸收利用的同时,可能会引起土壤磷库的变化,进而会对土壤细菌群落结构产生一定的影响。土壤微生物群落多样性与土壤功能密切相关,土壤环境的改变会直接引起微生物群落结构变化[10]。因此,磷高效转基因水稻在提高自身对土壤磷素高效吸收利用的同时,是否对土壤细菌多样性产生影响还不得而知。目前,有关磷高效转基因作物种植对土壤细菌多样性影响的报道较少。本研究主要利用PCR-DGGE方法,研究磷高效转基因水稻OsPT4的种植对土壤细菌群落多样性的影响,为该磷高效转基因水稻新材料将来的应用及其环境安全性评价提供理论依据。
1 材料和方法 1.1 试验地概况与试验设计试验地位于农业部环境保护科研监测所网室内,种植小区为长、宽、深均为1 m的水泥池,池内填充采自天津津南区的潮土,部分基本理化性质如下:全磷含量1.19 g·kg-1,全氮含量0.96 g·kg-1,有机质含量24.55 g·kg-1,pH8.21。
试验设不施磷和施磷15 g·m-2两个处理,各5次重复。磷源为KH2PO4,全部用作基肥。以尿素和硫酸钾作为N、K源,施氮20 g·m-2,其中50%作为基肥于种植前施用,50%作追肥施用,施钾18 g·m-2全部用作基肥。
1.2 供试材料试验所用水稻为磷高效转基因水稻OsPT4、磷高效突变体水稻PHO2及其非转基因亲本水稻材料日本晴,均由南京农业大学资源与环境科学学院植物营养分子生物学实验室提供。
1.3 土壤样品采集水稻种子于2014年5月20日播种在培养盘中,每孔5粒,于7月1日移苗,每个水泥池内移栽水稻30株。在水稻分蘖期、拔节期、抽穗扬花期和成熟期分别采集土样。采集时,去除表面杂草和枯枝落叶,分别在各水泥池内选取3株水稻,用直径3.5 cm的土钻在距水稻主茎2 cm处取20 cm深的土样,并将各个采样区的样品分别混合,鲜土样置于-20 ℃冰箱,用于土壤细菌群落多样性分析,另一部分经风干、研磨、过筛用于土壤理化性质的测定。
1.4 测定方法 1.4.1 土壤总DNA提取采用MoBio公司的Powerlyzer powersoil DNA isolation kit(MoBio laboratories,SolanaBeach,CA,USA)试剂盒,取0.5 g鲜土置于Glass Bead Tube中,按操作说明逐步进行提取,提取到土壤的DNA用1.0%的琼脂糖凝胶检测样品质量,提取的DNA于-20 ℃保存。
1.4.2 PCR扩增将土壤DNA采用细菌16S rDNA V3可变区通用引物341f-GC(5′-CCTACGGGAGGCAGCAG-3′)和534r(5′-ATTACCGCGGCTGCTGG-3′)进行PCR扩增[11],5′端GC夹序列CGCCCGCCGCGCGCGGCGGG-CGGGGCGGGGGCACGGGGGG。PCR反应体系为50 μL(两种引物各1.0 μL,Premix Ex Taq 25 μL,稀释2倍的土壤DNA模板1.0 μL,用灭菌水补足至50 μL)。PCR反应条件为:95 ℃预变性5 min;94 ℃变性1 min,57 ℃退火1 min,72 ℃延伸2 min,35个循环;72 ℃延伸5 min。PCR产物用1.5%的琼脂糖凝胶电泳进行检测。
1.4.3 DGGE检测和条带回收PCR产物采用Bio-Rad公司的DcodeTM通用突变检测系统(Bio-Rad,USA)按照操作说明进行检测。主要步骤如下:浓度为8%聚丙烯酰胺,变性梯度为40%~60%,60 ℃预热,将30 μL PCR产物与15 μL 6×loading buffer混合后用微量进样器加入胶孔,先在60 ℃、60 V恒定电压下预跑60 min,然后在60 ℃、100 V电泳12 h。电泳完毕后用SYBR R Green1(1:10 000)染色30 min,再用Gel Dox XR凝胶成像系统(Bio-Rad)进行观察与拍照。然后在紫外灯下对图谱的特异条带和优势条带进行割胶,切割回收的条带置于1.5 mL灭菌管内,加500 μL去离子水4 ℃过夜。
1.4.4 DGGE条带纯化和克隆回收处理过的条带用341f和534r引物进行扩增。将扩增产物割胶回收,用Wizad® SV Gel and PCR Clean-Up system试剂(Progema,USA)纯化,并与载体pGEM-T Easy Vector(Progema,USA)连接转化,挑取培养后的白色菌落,接种到液体培养基中,37 ℃摇床培养10 h。
1.4.5 测序及序列比对分别吸取各样品菌液1 mL,用T7和SP6两种引物进行测序(由上海生工生物工程技术服务有限公司完成)。将测序得到的DNA序列与NCBI数据库中已有的序列进行比对,获得相近的典型菌株序列。
1.5 数据分析采用SPSS 17.0(Duncan′s test)对试验数据进行分析,采用Quantity One 4.6.2软件进行数字化处理并进行聚类分析。各样品用香农-维纳指数(Shannon-Wiener index,H)、均匀度(Evenness index,EH)和丰富度(Richness,S)评价细菌多样性的变化,其计算公式如下:

对16S rDNA产物DGGE指纹图谱(图 1)分析表明:施磷处理时,在不同生长期内,磷高效转基因水稻OsPT4和非转基因水稻日本晴、PHO2的DGGE指纹图谱间有较大的相似性,多数为共有条带,表明这些条带代表的土壤细菌类群比较稳定,不受水稻品种的影响。OsPT4仅在抽穗扬花期与日本晴有1条差异条带(图 1C箭头所示),PHO2在拔节期与日本晴有1条差异条带(图 1B箭头所示)。

|
| 图 1 施磷处理不同生长期不同土壤样品DGGE 指纹图谱 Figure 1 DGGE profile in soil grown with different rice varieties under phosphate fertilizer application |
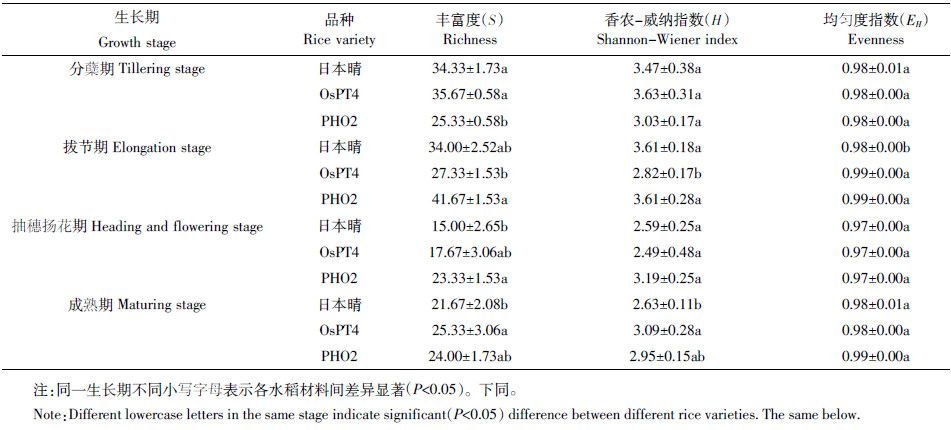
在施磷处理的不同生长期内,土壤细菌丰富度在各生长期内均呈现升-降-升的变化趋势,三个品种的土壤细菌丰富度在抽穗扬花期均为最低。OsPT4、PHO2在拔节期和成熟期的土壤细菌香农-维纳指数与日本晴相比具有显著差异,OsPT4与PHO2、日本晴的均匀度指数仅在拔节期具有显著差异(表 1)。
 |
不施磷条件下,对16S rDNA产物DGGE指纹图谱(图 2)分析表明:不同生长期内,OsPT4和日本晴、PHO2的DGGE指纹图谱有较大的相似性,多数条带为共有条带,表明这些条带代表的土壤细菌类群也比较稳定,也不受水稻品种的影响。但在分蘖期和成熟期出现了差异性条带。在水稻分蘖期,OsPT4与日本晴相比,增加三条差异条带(图 2B);在水稻成熟期,OsPT4与PHO2、日本晴相比,增加一条差异条带(图 2D)。

|
| 图 2 不施磷处理不同水稻生长期不同土壤样品DGGE 指纹图谱 Figure 2 DGGE profile in soil grown with different rice varieties without phosphate fertilizer application |
不施磷处理的水稻在各个生长期内,土壤细菌丰富度均呈现先升高后降低的趋势(表 2),三个水稻品种的丰富度指数均在水稻抽穗扬花期最低。OsPT4的香农-维纳指数与日本晴相比,在分蘖期、拔节期、成熟期显著增加;在抽穗扬花期OsPT4、PHO2的均匀度指数与日本晴相比显著增加。
 |
根据土壤细菌DGGE图谱数字化结果,分别选择施磷处理DGGE凝胶上主要条带36条(8~42)、不施磷处理DGGE凝胶上主要条带43条(a1~a43)和差异性条带6条(1~6)进行割胶纯化、连接转化后测序,条带位置如图 1、图 2所示。将6条特异性条带测序结果在NCBI数据库中进行Blast比对,将匹配度高的序列作为比对结果,如表 3所示,施磷处理时,PHO2在拔节期的差异条带为1(Uncultured Cyanobacterium),属于蓝藻菌门(Cyanobactteria)藻青菌属(Cyanobacterium);OsPT4在抽穗扬花期缺失的条带为条带2(Uncultured Gemmatimonadetes bacterium),属于芽单胞菌门(Gemmatimonadetes);不施磷处理时,磷高效转基因水稻OsPT4在分蘖期增加的条带自下而上依次为:条带3(Uncultured Geobacter sp.),属于变形菌门(Proteobacteria)地杆菌属(Geobacter);条带4(Bacillus sp.),属于厚壁菌门(Firmicutes)芽孢杆菌属(bacillus);条带5为(Uncultured Chloroflexi bacterium)属于绿弯菌门(Chloroflexi);OsPT4在水稻成熟期增加的条带为条带6(Uncultured Geoalkalibacter sp.),属于变形菌门(Proteobacteria)地杆菌属(Geobacter)。除条带4外,其余条带均属于不可培养微生物。
 |
转基因作物外源蛋白可以通过根系分泌物、作物残茬和花粉传播等多条途径进入土壤[12],可能对土壤微生物的种类、数量以及多样性产生一定影响。因此,随着转基因作物的大面积种植,转基因作物对土壤微生物多样性的潜在影响越来越受到关注。Kim等[13]使用PCR-DGGE方法对Iksan 483、Milyang 204两种转基因水稻和四种非转基因水稻进行研究,指出转基因水稻种植对土壤细菌群落没有显著影响,但是在8月到12月之间土壤细菌群落显示出季节性的差异。宋亚娜等[14]通过连续两年种植转cry1Ac/cpti双价抗虫基因水稻,发现一定时期内转基因水稻种植对土壤氨氧化细菌的群落组成和丰度没有显著影响。有些研究同样发现转基因作物种植对土壤微生物多样性不产生显著影响[15, 16],然而也有研究得出不同的结论。陈丽华等[17]通过室内秸秆还田模拟试验研究广谱抗真菌蛋白转基因水稻对土壤真菌群落的影响,指出在秸秆还田前期(40 d),转基因秸秆处理的土壤和非转基因秸秆处理的土壤其真菌群落结构存在显著差异,而在处理后期(50~90 d)则不存在差异,说明转基因水稻对土壤群落结构存在影响,虽然这种影响是短暂的。Castaldini等[18]在温室环境种植转Bt基因玉米和非转基因玉米,发现种植转Bt基因玉米对土壤微生物群落结构、分布及活性产生了显著影响。
本试验中,从香农-威纳指数来看,两种不同处理,在水稻生长的拔节期和成熟期,OsPT4与PHO2、日本晴之间都存在显著差异(P < 0.05);从均匀度指数来看,仅在施磷处理的拔节期和不施磷处理的抽穗扬花期,OsPT4、PHO2与对照日本晴之间存在显著差异(P < 0.05),其余时期未发现显著差异;从土壤细菌的丰富度来看,两种不同处理时,三种水稻的土壤细菌丰富度都呈现先降后升的趋势,且在抽穗扬花期土壤细菌丰富度最低,同一处理同一时期磷OsPT4的土壤细菌丰富度比日本晴高,但差异不显著,可能是由于磷高效转基因水稻在将土壤中更多的磷转化为植株可吸收利用形态的同时,也使得土壤中与磷形态有关的细菌类群增加。
综上所述,在水稻生长的不同时期,磷高效转基因水稻对土壤细菌多样性的影响也不同。这与陈晓雯等[19]的研究一致,她通过田间试验研究2种转基因水稻对土壤微生物群落结构的影响,指出仅在水稻生长旺盛期对土壤微生物群落有显著影响,在水稻抽穗期和成熟期则没有显著影响。金凌波等[20]也同样发现磷高效转基因大豆与非转基因大豆的土壤微生物数量和群落多样性存在一些差异,但这种因磷高效转基因大豆种植所产生的影响却不如生育期和季节变化对土壤微生物数量和群落产生的影响大。
从DGGE图谱可以看出三种水稻土壤细菌群落有很强的相似性,仅存在个别条带亮度不同及6条差异性条带。差异性条带克隆结果显示,OSPT4增加的4条差异条带分别为:地杆菌属(2条)、芽孢杆菌属(1条)、绿弯菌门(1条),说明在水稻生长旺盛期磷高效转基因水稻使得土壤中的地杆菌属、芽孢杆菌属以及绿弯菌门丰富度增加。地杆菌属属于δ-变形菌门(δ-Proteobacteria)地杆菌科(Geobacteraceae),是各种地下沉积环境的优势物种,它具有修复污染环境的能力[21, 22];芽孢杆菌属是最大的细菌属,包括276个种和7个亚种,它可以降解土壤中难溶的化合物,固定空气中的氮[23, 24];绿弯菌门普遍存在于各种自然环境和特定环境中,是有机质富集的地下水生物圈最大的细菌群,在生态系统中发挥不可或缺的作用[25]。施肥时,OsPT4缺失的条带属于芽单胞菌门,广泛存在于陆地生态系统,但其在土壤微生物群落中所占的比例仅为2%左右,并且因为缺乏合适的培养条件对其生态功能所知甚少[26]。同样,El-chakhtoura等[27]指出芽单胞菌门占微生物群落比例较小,不能确定影响其群落的具体因素。
影响土壤微生物群落多样性的因素有很多,如作物种类、根系分泌物、土壤类型及养分状况和耕作方式等[28],这些因素间往往存在相互作用,某一因素的改变通常会导致其他因素的改变,最终影响到土壤微生物的群落结构。因此,要研究磷高效转基因水稻对土壤细菌多样性的影响,还需要对多因素间的相互作用进行研究。本研究是通过室外小区试验进行的短期研究,得出磷高效转基因水稻的种植在个别生长期对土壤细菌群落多样性有影响,然而多数是未培养的土壤细菌。鉴于土壤细菌数量众多,且种类复杂,磷高效转基因水稻长期种植是否对土壤细菌群落多样性产生影响,还需要进行长期的田间试验进一步研究。
4 结论磷高效转基因水稻OsPT4各生长期的土壤细菌丰富度与PHO2、日本晴规律一致,均呈现先降后升的趋势,且最小值出现在水稻的抽穗扬花期;与日本晴、PHO2相比,OsPT4种植在水稻的拔节期、成熟期对香农-维纳指数存在显著影响;与日本晴相比,OsPT4、PHO2种植仅在施磷处理的拔节期和不施磷处理的抽穗扬花期对均匀度指数产生显著影响。总之,磷高效转基因水稻对土壤细菌多样性的影响因水稻生长期、施肥处理的不同而有所差异。
| [1] | James C. 2014年全球生物技术/转基因作物商业化发展态势[J]. 中国生物工程杂志, 2015, 35(1):1-14. James C. Global biotech/GM crops commercialization development situation in 2014[J]. China biotechnology, 2015, 35(1):1-14. |
| [2] | 卢荣宝, 夏辉. 转基因植物的环境生物安全:转基因逃逸及其潜在生态风险的研究和评价[J]. 生命科学, 2011, 23(2):186-194. LU Bao-rong, XIA Hui. Environmental biosafety of transgenic plants:Research and assessment of transgene escape and its potential ecological impacts[J]. Chinese Bullettin of Life Sciences, 2011, 23(2):186-194. |
| [3] | Neher D, Muthumbi A W N, Dively G P. Impact of coleopteran-active Bt corn on non-target nematodecommunities in soil and decomposing corn roots[J]. Soil Biology & Biochemistry, 2014, 76:127-135. |
| [4] | Tan F X, Wang J W, Chen Z N, et al. Assessment of the arbuscular mycorrhizal fungal community in roots and rhizosphere soils of Bt corn and their non-Bt isolines[J]. Soil Biology & Biochemistry, 2011, 43:2473-2479. |
| [5] | Roger A, Libohova Z, Rossier N, et al. Spatial variability of soil phosphorus in the Fribourgcanton, Switzerland[J]. Geoderma, 2014, 217-218:26-36. |
| [6] | Hu B, Zhu C G, Li F, et al. LEAF TIP NECROSIS1 plays a pivotal role in regulation of multiple phosphate starvation responses in rice[J]. Plant Physiology, 2011, 156(3):1101-1115. |
| [7] | Zhang F, Sun Y F, Pei W X, et al. Involvement of OsPht1;4 in phosphate acquisition and mobilization facilitates embryo development in rice[J]. The Plant Journal, 2015, 82:556-569. |
| [8] | 吴娜. 水稻磷转运蛋白OsPT4的生理功能鉴定[D]. 南京:南京农业大学, 2011. WU Na. Physilogical functional identification of rice phosphate transporter OSPT4[D]. Nanjing:Nanjing Agricultural University, 2011. |
| [9] | Aung K, Lin S I, Wu C C, et al. PHO2, a phosphate overaccumulator, is caused by a nonsense mutation in a MicroRNA399 Target Gene[J]. Plant Physiology, 2006, 141:1000-1011. |
| [10] | 张晶, 张惠文, 李新宇, 等. 土壤微生物生态过程与微生物功能基因多样性[J]. 应用生态学报, 2006, 17(6):1129-1132. ZHANG Jing, ZHANG Hui-wen, LI Xin-yu, et al. Soil microbial ecological process and microbial functional gene diversity[J]. Chinese Journal of Applied Ecology, 2006, 17(6):1129-1132. |
| [11] | Proietti P, Federici E, Fidati L, et al. Effects of amendment with oil mill waste and its derived-compost on soil chemical and microbiological characteristics and olive(Oleaeuropaea L.) productivity[J]. Agriculture, Ecosystems and Environment, 2015(207):51-60. |
| [12] | 刘文娟, 刘勇. 转Bt基因作物毒素蛋白对土壤生态的影响[J]. 中国测试, 2009, 35(6):92-96. LIU Wen-juan, LIU Yong. Impact of toxin proteins released from transgenic Bt crops on soil ecosystems[J]. China Measurement & Test, 2009, 35(6):92-96. |
| [13] | Kim M C, Ahn J H, Shin H C, et al. Molecular analysis of bacterial community structures in paddy soils for environmental risk assessment with two varieties of genetically modified Rice, Iksan 483 and Milyang 204[J]. Biotechnol, 2008, 18(2):207-218. |
| [14] | 宋亚娜, 苏军, 林艳, 等. 转cry1Ac/cpti基因水稻对土壤氨氧化细菌群落组成和丰度的影响[J]. 生物安全学报, 2012, 21(1):67-73. SONG Ya-na, SU Jun, LIN Yan, et al. Effect of cry1Ac/cpti transgenic rice on community composition and abundance of ammonia-oxidizing bacteria in paddy soil[J]. Journal of Biosafety, 2012, 21(1):67-73. |
| [15] | 李刚, 赵建宁, 杨殿林. 抗草甘膦转基因大豆对根际土壤细菌多样性的影响[J]. 中国农学通报, 2011, 27(1):100-104. LI Gang, ZHAO Jian-ning, YANG Dian-lin. Effects of glyphosate resistant transgenic soybean on bacterial diversity in rhizospheric soil[J]. Chinese Agricultural Science Bulletin, 2011, 27(1):100-104. |
| [16] | Fang H, Dong B, Yan H, et al. Effect of vegetation of transgenic Bt rice lines and their straw amendment on soil enzymes, respiration, functional diversity and community structure of soil microorganisms under field conditions[J]. Journal of Environmental Sciences, 2012, 24(7):1259-1270. |
| [17] | 陈丽华, 吕新, 林碧娇, 等. 广谱抗真菌蛋白转基因水稻秸秆模拟还田对土壤真菌群落结构的影响[J]. 中国生态农业学报, 2015, 23(1):87-94. CHEN Li-hua, LÜ Xin, LIN Bi-jiao, et al. Effects of simulated straw return of transgenic rice expressing broadspectrum antifungal proteins on soil fungal community structure[J]. Chinese Journal of Eco-Agriculture, 2015, 23(1):87-94. |
| [18] | Castaldini M, Turrini A, Sbrana C, et al. Impact of Bt corn on rhizospheric and soil eubacterial communities and on beneficial mycorrhizal symbiosis in experimental microcosms[J]. Applied and Environmental Microbiology, 2005, 71(11):6719-6729. |
| [19] | 陈晓雯, 林胜, 尤民生, 等. 转基因水稻对土壤微生物群落结构及功能的影响[J]. 生物安全学报, 2011, 20(2):151-159. CHEN Xiao-wen, LIN Sheng, YOU Min-sheng, et al. Effects of transgenic rice on the structure and function of soil microbial communities[J]. Journal of Biosafety, 2011, 20(2):151-159. |
| [20] | 金凌波, 周峰, 姚涓, 等. 磷高效转基因大豆对根际微生物群落的影响[J]. 生态学报, 2012, 32(7):2082-2090. JIN Ling-bo, ZHOU Feng, YAO Juan, et al. Effects of P-efficient transgenic soybean on rhizosphere microbial community[J]. Acta Ecologica Sinica, 2012, 32(7):2082-2090. |
| [21] | Tran H T, Krushka J, Antommattei F M, et al. Comparative genomics of Geobacter chemotaxis genes reveals diverse signaling function[J]. BMC Genomics, 2008, 9. doi:10.1186/1471-2164-9-471. |
| [22] | Aklujkar M, Young N D, Holmes D, et al. The genome of Geobacter bemidjiensis, exemplarfor the subsurface clade of Geobacter species that predominate in Fe(Ⅲ)-reducing subsurface environments[J]. BMC Genomics, 2010, 11. doi:10.1186/1471-2164-11-490. |
| [23] | Kim H J, Park C S, Lee S, et al. Bacillus cheonanensis sp. nov. isolated from near poultry farm soil[J]. Journal of Microbiology, 2014, 52(7):554-558. |
| [24] | 龚国淑, 唐志燕, 邓香洁, 等. 成都郊区土壤芽孢杆菌的空间分布及其多样性[J]. 生态学杂志, 2009, 28(10):2009-2013. GONG Guo-shu, TANG Zhi-yan, DENG Xiang-jie, et al. Spatial distribution and species diversity of soil Bacillus spp. in Chengdu suburbs[J]. Chinese Journal of Ecology, 2009, 28(10):2009-2013. |
| [25] | Yamada T, Sekiguchi Y. Cultivation of uncultured Chloroflexi subphyla:Significance and ecophysiology of formerly uncultured Chloroflexi‘subphylum i’with natural and biotechnological relevance[J]. Microbes Environ, 2009, 24(3):205-216. |
| [26] | DeBruyn J M, Nixon L T, Fawaz M N, et al. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil[J]. Applied and Environmental Microbiology, 2011, 77(17):6295-6300. |
| [27] | El-Chakhtoura J, Prest E, Saikaly P, et al. Dynamics of bacterial communities before and after distribution in a full-scale drinking water network[J]. Water Research, 2015, 74:180-190. |
| [28] | Dey R, Pal K K, Tilak K V B R. Influence of soil and plant types on diversity of rhizobacteria[J]. Proceedings of the National Academy ofSciences, 2012, 82(3):341-352. |