 2016, Vol. 35
2016, Vol. 35文章信息
- 蔡秋玲, 林大松, 王果, 王迪
- CAI Qiu-ling, LIN Da-song, WANG Guo, WANG Di
- 不同类型水稻镉富集与转运能力的差异分析
- Differences in cadmium accumulation and transfer capacity among different types of rice cultivars
- 农业环境科学学报, 2016, 35(6): 1028-1033
- Journal of Agro-Environment Science, 2016, 35(6): 1028-1033
- http://dx.doi.org/10.11654/jaes.2016.06.002
文章历史
- 收稿日期: 2016-01-19

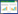




2. 农业部环境保护科研监测所, 天津 300191
2. Agro-Environmental Protection Institute, Ministry of Agriculture, Tianjin 300191, China
土壤重金属引起的农田污染及食品安全隐患[1],近几年成为备受关注的研究重点。水稻是我国重要的口粮作物,也是最易从土壤吸附重金属的作物之一。随着重金属在土壤中的不断累积及土壤酸化作用增强,水稻的重金属超标事件不断发生[2],特别是重金属Cd。不同品种水稻的Cd富集能力存在差异,有研究表明不同水稻品种对Cd吸收差异使得其籽粒Cd 的积累成倍增加[3],主要原因是有毒重金属能够通过必需营养元素的离子通道进入植物体内[4]。周歆[5]和张标金[6]等研究也证实不同类型的水稻籽粒Cd含量相差数倍。此外,Yu等[7]发现在不同Cd浓度胁迫下,同一品种水稻籽粒Cd含量存在差异。说明水稻籽粒Cd含量的差异由遗传特性和土壤Cd浓度共同影响。近年来,随着水稻Cd富集能力研究的不断深入,发现这种差异的表现与水稻的生物量没有必然联系。不同品种水稻根系耐Cd的能力有所不同[8],而且Cd从水稻根部向上转运的强度以及地上部向籽粒的输送速率也存在差异[9]。这些因素共同影响着水稻品种间Cd富集的差异性。通过研究者的不断努力,发现水稻中存在许多与金属转运相关的蛋白家族[10],在作物体内对Cd的迁移过程发挥了重要作用。
水稻籽粒Cd富集是一个复杂的生理过程。目前水稻吸收及迁移转化Cd的相关研究有很多,但在水稻各器官对籽粒Cd积累的贡献方面还没有系统的研究。因此,本研究选择在自然条件下种植84个水稻品种,筛选具有相似产量与Cd吸附特性的品种进行归类分组,研究不同类型水稻各器官Cd富集及各环节转运特征,探讨影响水稻籽粒Cd富集的重要因素,从而为高产低Cd水稻品种的筛选提供理论基础。
1 材料与方法 1.1 土壤背景本实验为小区试验,选点湖南省湘潭市水稻田。试验田常年种植双季稻,前作为早稻。土壤的基本理化性质所选指标均采用常规方法测定,测定结果见表 1。

|
参试品种为适合湖南省种植的晚稻品种,共84个,编号及具体水稻品种名称见表 2。
试验品种(30 d苗龄)于2014 年 7 月 23 日移栽至水稻试验田,随机区组排列,四周设不少于5行的保护行。小区面积为3.5 m×3.5 m,株距和行距均为20 cm。田间栽培管理按常规措施进行,所有小区保持一致,期间未发生影响产量的病虫害,水稻于11 月 21 日收获。
1.3.1 采集样品及测定各小区按照五点采样法各采集一个混合水稻样品和土壤。水稻植株先后用自来水、蒸馏水清洗,将植株分为根、茎、叶、稻谷部分。稻谷风干称量为W1,按农业部颁布《米质测定方法》(NY147—1988)出糙。所有采集样品110 ℃杀青后,70 ℃烘至恒重。然后粉碎,过60目筛。样品中Cd含量采用HNO3湿消解法,用ICP-MS(型号:Agilent 7500 Series)测定。以国家标准物质GBW10045(GSB-23)作为质控,保证数据真实性。土样经自然风干,去除杂质,粉碎过100目筛,测土壤Cd全量,用ICP-MS(同上)测定,以GSS8标准物质为质控。
1.3.2 产量测定分别收获试验田各小区水稻。将水稻用碾米机分为稻草与稻谷两部分,自然风干,稻谷称重为W2,记录。各小区稻谷总产量为:W=W1+W2。
1.4 数据处理富集系数(AFA-土)=A器官中重金属含量/土中重金属含量
转运系数(TFA-B)= B器官中重金属含量/A器官中该重金属含量
采用Microsoft Excel 2003进行数据整理,运用SPSS18.0进行系统聚类、方差及相关性分析。
2 结果与分析 2.1 水稻产量与Cd富集能力的相关性分析本试验田为中轻度Cd污染区,Cd对水稻各项生理指标及自然产量没有产生明显影响。84个水稻品种的稻米Cd含量范围为0.04~0.40 mg·kg-1,说明不同品种的水稻糙米Cd含量存在差异。为减少污染物空间差异性的影响,本研究对作物及相应根系土壤进行一对一采样。测得作物根系土壤Cd全量范围为0.39~0.73 mg·kg-1,但作物Cd含量与土壤含量不存在一一对应性。根据糙米及土壤Cd含量计算每个品种的糙米Cd富集系数,将其作为反映Cd富集能力的指标。计算得Cd富集系数范围为0.10~0.78,平均值为0.28±0.13,水稻的产量范围为8.20~11.50 kg (以小区3.5 m×3.5 m计),平均产量为(9.80±0.51) kg。但水稻产量与糙米Cd富集系数之间不存在显著相关性(图 1)。

|
| 图 1 水稻糙米Cd富集系数与产量的相关性 Figure 1 Correlations between production and Cd accumulation factor of brown rice |
为进一步研究具有类似产量与Cd富集能力的水稻富集及转运特征,选取产量和Cd富集系数两个变量,按照最远邻元素分析的系统聚类方式,将所有水稻品种分为四组进行统计分析,水稻组别为高产高Cd(组1)、低产高Cd(组2)、高产低Cd(组3)和低产低Cd(组4)。分组后,通过对这两个变量进行新复极差比较,验证了组间具有显著差异(见表 3)。
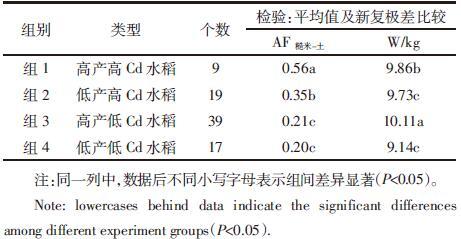
分组统计水稻各器官的富集系数。从图 2可知:四组数据中,不同类型水稻植株总体对Cd的富集能力顺序均为根系>茎>叶>糙米,产量和糙米Cd富集能力并不影响这一规律性。进入植株中的Cd主要在根部富集,差异性主要在于高Cd或高产水稻的各器官Cd富集能力均高于低Cd水稻。
探讨产量和Cd富集能力两个因子对水稻各器官富集系数的影响作用,其分析结果如图 2所示。当富集能力相似时(图 2a和2b),除了高产高Cd水稻的茎和糙米富集系数显著高于低产高Cd水稻,产量高低没有对各器官的富集系数产生明显影响。同为高产水稻时(图 2c),高Cd富集品种的茎、糙米富集系数显著高于相应的低Cd品种;而同为低产水稻时(图 2d),高Cd水稻各器官的富集系数显著高于相应的低Cd水稻。说明糙米Cd富集能力对水稻各器官富集系数影响要比产量的影响重要得多,茎和叶的富集系数更容易受到富集量和产量的影响。

|
| 图 2 不同产量及Cd富集能力的水稻组富集系数差异性比较 Figure 2 Comparison of Cd accumulation factor in different organs for various groups of rice with different production and Cd accumulation |
计算水稻各部位的Cd转运系数,统计每个水稻组数据。探讨产量和Cd富集系数两个因子对水稻各器官转运系数的影响作用,其分析结果如图 3所示。对于低Cd品种来说,产量没有对任何环节的转运系数产生明显影响,但对于高Cd(或高产)品种,高产(高Cd)品种根-茎、茎-糙米和叶-糙米的转运系数显著高于低产(或低Cd)品种;而同为低产品种时,高Cd富集品种的土-根、根-茎转运系数显著高于低Cd品种。说明高产或高Cd 品种有较强的将Cd从根转运到茎和从茎、叶转运到米的能力,而相对于低产低Cd品种来说,低产高Cd品种则有相对较强的将Cd 从土壤转运至根系、根系转运至茎的能力,但在继续向上转运的能力不明显。

|
| 图 3 不同产量及Cd富集能力的水稻组转运系数差异性比较 Figure 3 Comparison of Cd transfer factor among different organs for various groups of rice with different production and Cd accumulation |
Cd在土壤-植物系统中的迁移与分配是多重因素共同决定的。水稻籽粒Cd含量主要由根系的吸收及向地上部转运决定的。根系是Cd进入水稻体内的首要门户,是Cd的主要积累器官[11]。Nocito等[12]研究表明根对进入植物体中的Cd富集能力在49%~79%之间,潜在移动的Cd离子约为总Cd的24%。进入植物根部的Cd一部分累积在根系细胞壁和液泡中[13],一部分通过质外体和共质体这两种途径进行运输[14]。通过对比各组水稻根Cd富集系数发现,除了低产高Cd水稻和低产低Cd水稻的Cd富集系数之间差异显著,其他两组对比,均无显著差异。表明根对Cd的吸收并不能直接反映籽粒镉积累状况,与胡莹等[15]研究结果一致。
地上部的转运主要经历的过程包括了Cd离子进入木质部、木质部的转运及韧皮部的输送。根中Cd离子通过内皮层并入共质体途径,进入中柱的木质部才能继续向上转运,并在叶片和茎秆中积累[16]。当水稻进入灌浆期,茎节的Cd优先输送到谷粒,叶面的Cd再转运到茎节也输送到谷粒中。Uraguchi等[17]研究发现水稻木质部的Cd浓度与地上部包括籽粒Cd含量呈显著相关。Liu等[18]研究发现籽粒Cd含量与Cd从茎叶向籽粒转运能力呈显著正相关。Tanaka等[19]研究发现90%的Cd通过韧皮部输送实现水稻籽粒的积累,说明韧皮部的输送速率决定着籽粒的Cd积累水平。根系吸收的Cd一部分通过穗轴直接进入籽粒,另一部分转运到叶片储存起来,在灌浆期通过穗轴进入籽粒[20]。Rodda等[21]研究表明糙米中60%的Cd含量是由剑叶、茎秆等在水稻开花灌浆前累积的Cd重新活化,通过韧皮部输入籽粒。在水稻组间比较分析中发现,除了低产低Cd水稻与高产低Cd水稻的茎和米Cd富集系数无显著差异,其他两两组对比,均有显著差异。说明籽粒Cd的富集,茎叶起了直接作用,是籽粒积累Cd的通路之一,与居学海等[22]研究结果一致。
随着对水稻Cd吸收转运分子机制的深入探究,目前已发现一些与Cd转运相关的基因对地上部的转运发挥了巨大作用。而很多基因的存在不仅仅是控制水稻Cd的通路,同时具有影响多种元素共吸收的能力。如Takahashi[23-24]研究发现OsHMA2基因参与地下部向地上部的转运,对重金属Cd和Zn在木质部装载都发挥作用。Uraguchi等[25-26]研究发现,在茎叶及节点存在一些与Cd吸收相关基因,对籽粒Cd的富集起到重要作用。如OsLCT1基因主要在水稻生殖阶段的叶片和水稻茎节高强度表达,通过韧皮部参与Cd向谷粒运输过程。这也充分证明,地上部对水稻籽粒Cd的富集起到重要作用。从基因层面探究将是未来研究的重点。
从本文研究结果可见,产量对于水稻Cd吸收有一定影响,但是最重要的影响因素还是水稻本身富集能力,一些水稻品种存在保持低Cd富集基因。因此,筛选、培育适合在中轻度污染区种植的高产低Cd水稻品种是可行的。本研究84个品种中,筛选出39个高产低Cd水稻品种。此外,在水稻组间比较分析发现,水稻的茎叶是影响其重金属吸收的重要环节,由此在水稻生产过程中配合控制根的吸收,辅以阻隔重金属在茎、叶环节的转移等手段,例如延长淹水时间、喷施叶面调理剂,都有可能降低稻米超标的潜在风险。
4 结论(1) 从84个水稻品种统计结果可见,水稻产量与糙米Cd富集能力不存在相关性。按产量和糙米Cd富集能力两指标划分的水稻组别中,各组水稻各器官Cd富集能力和Cd在水稻体内各环节的转运能力存在差异。
(2) 不同类型水稻中各器官Cd富集规律均为根系>茎>叶>糙米,主要差异在于茎-叶的富集与转运,这可能是控制水稻籽粒Cd积累的重要因素。
(3) 筛选、培育适合在中轻度污染区种植的高产低Cd水稻品种是可行的。在种植过程中控制茎叶的吸收与转运可降低水稻籽粒重金属Cd超标风险。
| [1] | 曾希柏, 徐建明, 黄巧云, 等. 中国农田重金属问题的若干思考[J]. 土壤学报, 2013, 50 (1) : 186–194 . ZENG Xi-bai, XU Jian-ming, HUANG Qiao-yun, et al. Some deliberations on the issues of heavy metals in farmlands of China[J]. Acta Pedologica Sinica, 2013, 50 (1) :186–194 . |
| [2] | 詹杰, 魏树和, 牛荣成. 我国稻田土壤镉污染现状及安全生产新措施[J]. 农业环境科学学报, 2012, 31 (7) : 1257–1263 . ZHAN Jie, WEI Shu-he, NIU Rong-cheng. Advances of cadmium contaminated paddy soil research and new measure of its production in China:A review[J]. Journal of Agro-Environment Science, 2012, 31 (7) :1257–1263 . |
| [3] | 史静, 潘根兴, 张乃明. 镉胁迫对不同杂交水稻品种Cd、Zn吸收与积累的影响[J]. 环境科学学报, 2013, 33 (10) : 2904–2910 . SHI Jing, PAN Gen-xing, ZHANG Nai-ming. Effect of cadmium stress on Cd and Zn uptake and accumulation of different cultivars of hybrid rice[J]. Acta Scientiae Circustantiae, 2013, 33 (10) :2904–2910 . |
| [4] | Bernal M, Testillano P, Alfonso M, et al. Identification and subcellular localization of the soybean copper P1B-ATPase GmHMA8 transporter[J]. Journal of Structural Biology, 2007, 158 (1) :46–58 . |
| [5] | 周歆, 周航, 胡淼, 等. 不同杂交水稻品种糙米中重金属Cd、Zn、As含量的差异研究[J]. 中国农学通报, 2013, 29 (11) : 145–150 . ZHOU Xin, ZHOU Hang, HU Miao, et al. The difference of Cd, Zn and As accumulation in different hybrid rice cultivars[J]. Chinese Agricultural Science Bulletin, 2013, 29 (11) :145–150 . |
| [6] | 张标金, 罗林广, 魏益华, 等. 不同基因型水稻镉积累动态差异分析[J]. 中国农学通报, 2015, 31 (9) : 25–30 . ZHANG Biao-jin, LUO Lin-guang, WEI Yi-hua, et al. Analysis of cadmium accumulation dynamics in rice with distinct genotypes[J]. Chinese Agricultural Science Bulletin, 2015, 31 (9) :25–30 . |
| [7] | Yu H, Wang J L, Fang W, et al. Cadmium accumulation in different rice cultivars and screening for pollution-safe cultivars of rice[J]. Science of the Total Environment, 2006, 370 (2/3) :302–309 . |
| [8] | 何俊瑜, 任艳芳, 王阳阳, 等. 不同耐性水稻幼苗根系对镉胁迫的形态及生理响应[J]. 生态学报, 2011, 31 (2) : 522–528 . HE Jun-yu, REN Yan-fang, WANG Yang-yang, et al. Root morphological and physiological responses of rice seedlings with different tolerance to cadmium stress[J]. Acta Ecologica Sinica, 2011, 31 (2) :522–528 . |
| [9] | 龙小林, 向珣朝, 徐艳芳, 等. 镉胁迫下籼稻和粳稻对镉的吸收、转移和分配研究[J]. 中国水稻科学, 2014, 28 (2) : 177–184 . LONG Xiao-lin, XIONG Xun-chao, XU Yan-fang, et al. Absorption, transfer and distribution of Cd in Indica and Japonica rice under Cd stress[J]. Chinese Journal of Rice Science, 2014, 28 (2) :177–184 . |
| [10] | 杨菲, 唐明凤, 朱玉兴. 水稻对镉的吸收和转运的分子机理[J]. 杂交水稻, 2015, 30 (3) : 2–8 . YANG Fei, TANG Ming-feng, ZHU Yu-xing. Molecular mechanism of cadmium absorption and transport in rice[J]. Hybrid rice, 2015, 30 (3) :2–8 . |
| [11] | 肖美秀, 林文雄, 陈祥旭, 等. 镉在水稻体内的分配规律与水稻镉耐性的关系[J]. 中国农学通报, 2006, 22 (2) : 379–381 . XIAO Mei-xiu, LIN Wen-xiong, CHEN Xiang-xu, et al. The relation between the law of Cd distribution in rice and the Cd-tolerance[J]. Chinese Agricultural Science Bulletin, 2006, 22 (2) :379–381 . |
| [12] | Nocito F, Lancilli C, Dendena B, et al. Cadmium retention in rice roots is influenced by cadmium availability, chelation and translocation[J]. Plant Cell and Environment, 2011, 4 (6) :994–1008 . |
| [13] | 吴倩. 不同Cd水平对水稻材料根系形态及Cd亚细胞分布的影响[D]. 成都:四川农业大学, 2009:13-19. WU-Qian. The effect of different cadmium levels on morphology and subcellular distribution in rice root[D]. Chengdu:Sichuan Agricultural University, 2009:13-19. |
| [14] | Ueno D, Koyama E, Yamaji N, et al. Physiological, genetic, and molecular characterization of a high-Cd-accumulating rice cultivar, Jarjan[J]. Experimental Botany, 2011, 62 (7) :2265–2272 . |
| [15] | 胡莹, 黄益宗, 段桂兰, 等. 镉对不同生态型水稻的毒性及其在水稻体内迁移转运[J]. 生态毒理学报, 2012, 7 (6) : 664–670 . HU Ying, HUANG Yi-zong, DUAN Gui-lan, et al. Cadmium toxicity and its translocation in two ecotype rice cultivars[J]. Asian Journal of Ecotoxicology, 2012, 7 (6) :664–670 . |
| [16] | Uranguchi S, Fujiwara T. Rice breaks ground for cadmium-free cereals[J]. Plant Biology, 2013, 16 (3) :328–334 . |
| [17] | Uraguchi S, Mori S, Kuramata M, et al. Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice[J]. Journal of Experimental Botany, 2009, 60 (9) :2677–2688 . |
| [18] | Liu J G, Qian M, Cai G L, et al. Uptake and translocation of Cd in different rice cultivars and the relation with Cd accumulation in rice grain[J]. Journal of Hazardous Materials, 2007, 143 (1/2) :443–447 . |
| [19] | Tanaka K, Fujimaki S, Fujiwara T, et al. Cadmium concentrations in the phloem sap of rice plants(Oryza saliva L.) treated with a nutrient solution containing cadmium[J]. Soil Science and Plant Nutrition, 2003, 49 (2) :311–313 . |
| [20] | 文志琦, 赵艳玲, 崔冠男, 等. 水稻营养器官镉积累特性对稻米镉含量的影响[J]. 植物生理学报, 2015, 51 (8) : 1280–1286 . WEN Zhi-qi, ZHAO Yan-ling, CUI Guan-nan, et al. Effects of cadmium accumulation characteristics in vegetative organs on cadmium content in grains of rice[J]. Plant Physiology Journal, 2015, 51 (8) :1280–1286 . |
| [21] | Rodda M S, Li G, Reid R J. The timing of grain Cd accumulation in rice plants:the relative importance of remobilization within the plant and root Cd uptake post-flowering[J]. Plant and Soil, 2011, 347 (1/2) :105–114 . |
| [22] | 居学海, 张长波, 宋正国, 等. 水稻籽粒发育过程中各器官镉积累量的变化及其与基因型和土壤镉水平的关系[J]. 植物生理学报, 2014, 50 (5) : 634–640 . JU Xue-hai, Zhang Chang-bo, SONG Zheng-guo, et al. Changs in cadmium accumulation in rice organs during grain development and their relationship with genotype and cadmium levels in soil[J]. Plant Physiology Journal, 2014, 50 (5) :634–640 . |
| [23] | Takahashi R, Ishimaru Y, Shimo H, et al. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice[J]. Plant Cell and Environment, 2012, 35 (11) :1948–1957 . |
| [24] | Yamaji N, Xia J X, Ueno M N, et al. Preferential delivery of zinc to developing tissues in rice is mediated by P-Type heavy metal ATPase OsHMA2[J]. Plant Physiology, 2013, 162 (2) :927–939 . |
| [25] | Uraguchi S, Kamiya T, Sakamoto T, et al. Low-affinity cation transporter(OsLCT1) regulates cadmium transport into rice grains[J]. Proc Natl Acad Sci USA, 2011, 108 (52) :20959–20964 . |
| [26] | Uraguchi S, Kamiya T, Clemens S, et al. Characterization of OsLCT1, a cadmium transporter from indica rice(Oryza sativa)[J]. Physiologia Plantarum, 2014, 151 (3) :339–347 . |