 2016, Vol. 35
2016, Vol. 35文章信息
- 陈惠君, 谭玲, 李取生, 罗涛, 方皓, 余丹萍, 胡妮, 杨耀帅
- CHEN Hui-jun, TAN Ling, LI Qu-sheng, LUO Tao, FANG Hao, YU Dan-ping, HU Ni, YANG Yao-shuai
- Cr/Pb低累积菜心品种筛选及其根际机理研究
- Screening and preliminary rhizosphere mechanisms of low Cr/Pb accumulation cultivars of Chinese flowering cabbages (Brassica parachinensis L.)
- 农业环境科学学报, 2016, 35(7): 1249-1256
- Journal of Agro-Environment Science, 2016, 35(7): 1249-1256
- http://dx.doi.org/10.11654/jaes.2016.07.004
文章历史
- 收稿日期: 2016-03-10






2. 广东省环境污染控制与修复材料中心, 广州 510632
2. Guangdong provincial research center of environmental pollution control and remediation material, Guangzhou 510632, China
铬(Cr)、铅(Pb)作为作物安全品质指标,通过食物链危及人类健康[1]。利用作物对重金属吸收累积的品种间差异,筛选、种植重金属低累积的农作物品种,已成为应对土壤重金属污染、保障食品安全的有效策略之一[2-4]。研究者多关注单一污染源下作物的低累积品种筛选[5-6],但土壤污染往往呈多种重金属复合污染。郭晓方等[7]研究通过田间实验发现灵丹20、正丹958 和高优1 号属Cd、Pb、Zn 和Cu 低累积玉米品种。目前多种重金属低累积品种的筛选研究还较少见诸报道[8]。
根系通过分泌有机物活化重金属,增加其生物有效性,从而促进根系对重金属的吸收[9]。重金属Cr、Pb作为植物非必需元素,可借用营养元素专一性离子通道进入根细胞内[10]。Ca 是植物所需大量营养元素,与Cr、Pb 皆为二价金属。有研究发现Cd、Pb 可能通过芦蒿质膜上的钙离子通道进入细胞内[11],作物低累积重金属的特性与吸收重金属途径[11]、根际分泌有机物[9]、根-茎转运[6]等相关。刘维涛等[12]研究表明15 种大白菜从土壤中吸收的Pb 主要集中和固定在根部,对可食部分累积Pb 有很大的影响。龚玉莲等[4]通过根箱试验发现蕹菜低Cd 品种的累积机制是由于其对土壤重金属活化能力较弱,从而降低对重金属的吸收累积。上述机制研究都在单一污染源下进行,而作物低累积重金属的根际机制在复合污染水平下研究较少。He 等[6]报道空心菜对多种重金属的累积差异是因为其根系分泌物存在品种差异,导致对根际土壤重金属活化不同。研究作物品种低累积多种重金属的机制有助于消减和抑制作物体内的重金属含量。
菜心(Brassicaparachinensis L.)是十字花科芸薹属蔬菜,是华南种植规模最大的叶菜类蔬菜作物之一[13]。已有研究表明芸薹属植物吸收重金属能力较强[8],故筛选、种植低累积重金属的菜心品种有助于降低长期食用菜心所带来的健康风险。本研究采集长期受Cr、Pb 复合污染的农田土壤进行盆栽实验,分析了37 个菜心品种的重金属Cr、Pb 累积能力差异,筛选出Cr/Pb 相对高累积和相对低累积的菜心品种,并进一步揭示品种低累积Cr、Pb 的根际机制,以期为保障农产品安全提供科学依据。
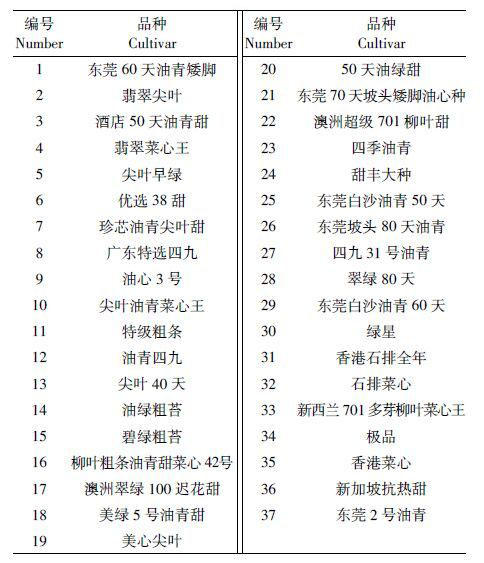
1 材料与方法 1.1 实验材料 1.1.1 供试菜心品种菜心种子共37 种,购自广州各农贸市场以及农科院种子市场,播种前于4 ℃保存。供试菜心名称及其编号列于表 1。
在广州郊区水稻田,采集耕作层0~20 cm 土壤,去掉颗粒大的杂质,粉碎,自然风干,过筛备用。供试土壤基本理化性质[14]:pH7.09,Pb 总量103.2 mg·kg-1,Cr 总量148.9 mg·kg-1,有机碳23.70 g·kg-1,阳离子交换量19.75 cmol·kg-1,全氮3.42 g·kg-1,全磷(以P 计)2.05 g·kg-1,全钾(以K 计)11.21 g·kg-1,碱解氮217mg·kg-1,速效磷(以P 计)127.8 mg·kg-1,速效钾(以K计)98.4 mg·kg-1。根据《食用农产品产地环境质量评价标准》HJ332—2006[15]蔬菜地的Cr、Pb 最高限值分别为150 mg·kg-1和50 mg·kg-1,供试土壤Pb 总量严重超标,是HJ 332—2006限值的2倍,Cr虽未超标,但大幅超过广东省土壤环境质量背景值50.5 mg·kg-1 [16],而且种植菜心品种亦出现部分样品Cr 含量超过食品安全标准现象。这表明,该供试土壤确实存在Cr、Pb复合污染。
1.2 实验设计 1.2.1 盆栽实验冬季在暨南大学温室中进行盆栽实验,保持温度在15~25 ℃。准确称2.2 kg 供试土壤置于高20 cm、直径5 cm 的塑料盆中。种植37 个菜心品种,每品种3个平行,一共111 盆,每盆6 株苗。将多菌灵浸泡2 h的种子播于塑料盆,覆土,早晚各浇灌自来水一次,50d 后收获植物。采用抖根法获得根际土[10],非根际土为远离根系生长区的土壤,用差速离心法离心:高速6000 r·min-1离心20 min,低速4000 r·min-1 离心20min,取上清液[12],用0.45 μm 孔径的抽滤膜进行抽滤,用石墨炉原子吸收分光光度计(岛津AA 7000)测定Cr、Pb 含量,用TOC 仪(岛津TOC-VCSH)测定溶解性有机碳(DOC)。
1.2.2 水培实验筛选相对高累积和相对低累积品种进行水培试验。种子消毒后,播种于水洗石英砂、覆砂,待菜心长至10 cm 时,挑选长势一致的苗,移至高15 cm、直径15cm 的1.5 L塑料桶中,每盆6 株,加入Hoagland 完全营养液[10],定时通气,待菜心长至4叶,更换含2 μmol·L-1Pb(NO3)2、2 μmol·L-1Cr(NO3)3·9H2O 的Hoagland完全培养液,用1 mol·L-1的HNO3或NaOH溶液将pH调至6.0,每隔4d更换培养液,30d后收获植物。
1.2.3 钙离子通道抑制剂实验随机挑选一个相对高累积和一个相对低累积品种,未添加抑制剂为对照组,添加抑制剂为处理组,每组每个品种3 个平行。在最后一次更换营养液12 h后,在对照组营养液中加入20 μmol·L-1Pb(NO3)2 和100 μmol·L-1Cr(NO3)3·9H2O,在处理组中加入相同量的重金属和1 mmol·L-1 钙离子通道抑制剂(LaCl3),处理8 h后收获植物。因为该实验为破坏性实验,钙离子通道抑制剂将抑制钙吸收,影响作物正常生长,所以处理时间必须较短。为了在短时间内观察到菜心体内重金属含量有无变化,实验按照Mei 等的方法[10],采用的重金属含量大幅高于根际土壤溶液实际含量。采收植物后,将植物根系剪下,双蒸水冲洗3 次,以滤纸吸干根系表面水分,将根系剪碎置于50 mL 离心管中,加入10 mL 浓度为10 mmol·L-1的EDTA-(NH4)2,放入超声仪中解析10 min,将根细胞壁中的离子解析出来[10]。此操作重复3次,将解析液转移至50 mL容量瓶,定容至50 mL,用于测定根系对重金属的吸附量。剩余部分根系在70 ℃烘箱中烘至恒重,称量干重并消解,用于测定根系对重金属的吸收量。
1.3 样品处理及测定清洗植物可食部分和根,置于滤纸上吸干植物表面水分,立即称鲜重,105 ℃杀青30 min,70 ℃烘至恒重,粉碎过筛后保存。称取0.4 g植物干样,采用硝酸+盐酸+氢氟酸方法微波消解,用原子吸收分光光度计(岛津AA 7000)测定Cr、Pb 含量,以国家标准物质GBW07602(GSV-1)为内标控制分析质量,同时做一份空白实验。
1.4 数据处理用Origin8.0 软件进行图形处理,SPSS 19.0 进行单因素方差分析、Duncun 多重比较。测试所得菜心可食部分和根部的重金属含量均为鲜重。转运系数为可食部分重金属含量与根部重金属含量之比,表示菜心根部重金属转运至可食用部分的能力。
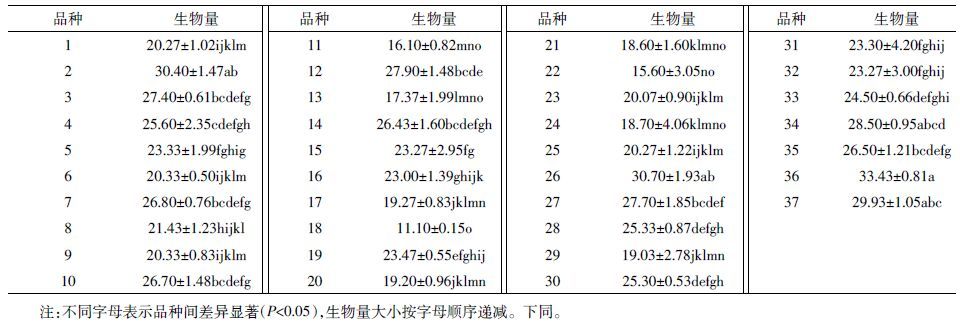
2 结果 2.1 盆栽条件下Cr/Pb 低累积菜心品种的筛选供试重金属污染水平下菜心未出现叶子发黄等不良症状。由表 2 可知,37 种菜心可食用部分生物量鲜重范围为14.5~32.4 g·pot-1。菜心生物量最大值与最小值之比为3倍,品种间生物量比值较大。这表明菜心生物量存在品种间差异,且对Cr、Pb 具有一定的耐受性。
|
菜心可食部分Cr、Pb 鲜重含量范围分别为0.151~0.630 mg·kg-1和10.38~24.34 μg·kg-1(图 1)。翡翠尖叶(2 号)、特级粗条(11 号)、油青四九(12 号)、澳洲超级701 柳叶(22 号)、四九31 号油青(27 号)菜心可食用部分Cr 含量超过《食品安全国家标准食品中污染物限量》GB 2762—2012[17]对Cr 的限值,长期食用具有一定的健康风险。而污染土壤Pb 总量虽是蔬菜产地最高限值[15]的2 倍,但菜心可食用部分Pb平均含量仅为GB 2762—2012[17]Pb 限值的5.3%,菜心受Pb 污染风险相对较小。

|
| 图 1 盆栽条件下菜心可食用部分Cr、Pb 含量 Figure 1 Cr and Pb concentrations in edible parts of the Chinese flowering cabbages in pot experiment GB 2762—2012 表示《食品安全国家标准食品中污染物限量》(Cr≤0.5 mg·kg-1、Pb≤0.3 mg·kg-1) |
在本实验中,Cr、Pb 可食用部分含量最大值与最小值之比分别为4.2 倍与2.4 倍,满足Cr/Pb 高累积和低累积品种筛选的需要[12, 18]。由图 2 可知,在供试土壤的Cr、Pb 复合污染条件下特级粗条、油青四九和四九31 号油青(11 号、12 号和27 号)所代表的点散落在直线上方附近,表示其同时吸收Cr、Pb 能力较强,属于Cr/Pb 高累积品种;甜丰大种、绿星、香港石排和极品(24号、30号、31号和34号)所代表的点散落在直线下方附近,表示其同时吸收Cr、Pb 能力较弱,属于Cr/Pb 低累积品种,且Cr、Pb 含量在高累积与低累积品种之间存在显著差异(P <0.05)。挑选Cr、Pb 含量较高的特级粗条、油青四九为Cr/Pb 相对高累积品种(Relativelyhigh Cr/Pb cultivars,HC),含量较低的绿星和极品为Cr/Pb 相对低累积品种(RelativelylowCr/Pbcultivars,LC)。

|
| 图 2 盆栽条件下菜心可食用部分Cr、Pb 含量的关系 Figure 2 Relation between Cr and Pb concentrations in edible parts of the Chinese flowering cabbages in potexperiment 盆栽条件下37 种菜心可食部分Cr质量浓度的对数 |
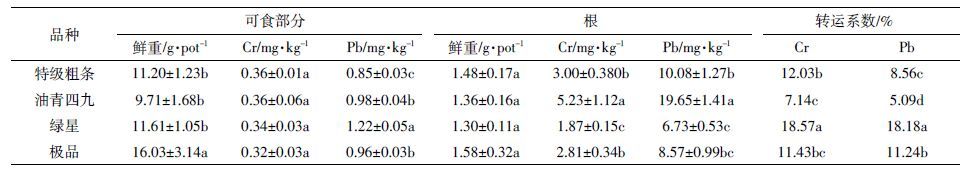
表 3 单因素方差分析结果表明,HC 可食部分Cr、Pb 含量显著高于LC,其根部Cr、Pb 含量也显著高于LC(P <0.05)。在可食部分和根部,菜心Cr、Pb 含量的品种差异表现一致(P <0.05)。HC 的特级粗条、油青四九根到可食部分Cr 转运系数显著高于LC 的绿星、极品,而Pb 的转运系数在HC/LC 之间无显著差异。

|
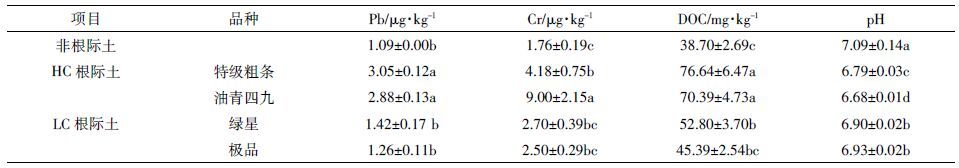
由表 4 可知,HC 的特级粗条、油青四九根际溶液Cr、Pb 含量,高于LC 的绿星、极品。供试土壤Cr、Pb总量较大但两种重金属在非根际土壤溶液中溶解态含量较低,说明作为已经被污染多年的土壤,其中重金属已经老化,生物有效性较低,菜心需活化土壤中的Cr、Pb 才能将其吸收进入体内。表 4 显示,与非根际土溶液相比,在根际溶液中Cr、Pb、DOC 均有不同的增幅,且LC 显著低于HC;根际土pH 均显著低于非根际土,且HC 的pH 值显著低于LC(P <0.05)。这说明与HC 相比,LC 分泌有机物与H+数量较少,活化土壤Cr、Pb 能力较低,导致土壤溶解态Cr、Pb 较少。
|
由表 5 可知,除了极品的可食部分生物量,水培条件下菜心各部分生物量不存在显著的品种差异。部分HC 根部Cr、Pb 含量显著高于LC(P <0.05),其中油青四九根部Cr、Pb 含量最高,绿星根部Cr、Pb 含量最低。根部Cr、Pb 含量的品种差异在盆栽和水培条件下表现基本一致。可食部分菜心的Cr、Pb 含量范围分别为0.32~0.36 mg·kg-1 和0.85~1.22 mg·kg-1,品种差异不显著。Pb 转运系数LC 显著高于HC,Cr 转运系数在HC/LC 之间只有绿星和油青四九有显著差异。
|
由表 6 可知,根鲜重不存在显著的品种差异和处理间差异(P <0.05)。由于水培溶液中Cr、Pb 的处理浓度较高,导致对照条件和添加抑制剂处理下,菜心品种的Cr 含量不存在显著的品种差异(P <0.05),Pb 含量仅在对照条件下存在显著品种差异(P <0.05),而Ca含量在对照和处理条件下仍然都存在显著品种差异(P <0.05)。在对照组,HC 根系对Ca 的吸收高于LC,证明LC 的钙离子通道较不活跃。钙离子通道抑制剂处理后特级粗条、绿星根部的Ca含量比对照组分别降低了34%、43%。与对照组相比,钙离子通道抑制剂使特级粗条(HC)的Cr、Pb 含量分别降低了60%、81%,使绿星(LC)的Cr、Pb含量分别降低了46%、87%。与对照组相比,在添加抑制剂处理组菜心根系的Ca、Cr、Pb含量显著下降,证明Cr、Pb 能通过钙离子通道进入根细胞内。
|
|
相对高累积品种可食部分Cr、Pb 含量均显著高于相对低累积品种(P <0.05,表 3)。植物吸收重金属并将其转运累积到地上部分需经一系列过程,包括植物活化根际土壤重金属、根细胞表面吸附以及跨质膜吸收、根系组织内的运输等[19],其中根际活化过程起重要作用。与非根际土溶液相比,HC/LC 根际土溶液中的Cr、Pb、DOC 含量较高,pH 较低,表明菜心根系分泌的有机物和H+对活化土壤Cr、Pb 有影响。在根际溶液中LC 的Cr、Pb、DOC 含量和根际土pH 均低于HC(表 4),表明LC 品种根系分泌有机物以及H+较少,对被土壤固定的Cr、Pb 活化能力较弱,部分导致了菜心可食部分Cr、Pb 的高/低累积差异。植物在正常生长情况下或受到重金属胁迫时都会分泌大量有机酸、氨基酸、糖类等物质,这些物质构成土壤DOC 的成分主体,DOC 与H+对植物活化土壤重金属具有调控作用,而分泌物的种类以及数量存在品种差异[20]。本研究中HC 的特级粗条、油青四九根际溶液DOC 和H+含量显著高于LC 的绿星、极品(表 4),导致HC 活化的根际土壤Cr、Pb 显著增加。龚玉莲等[4]研究也表明,与蕹菜低Cd 品种(QLQ)相比,H+在高Cd 品种(T308)的根际土壤中数量更多。He 等[6]亦报道了空心菜根系分泌物存在品种差异,导致根际土壤重金属活化也存在品种差异,从而引起了对多种重金属累积的品种差异。
3.2 Pb、Cr 同步高/低累积品种根部吸收、转运重金属能力的差异重金属Cr、Pb 作为植物非必需元素不具有专一的跨根系质膜离子通道。钙离子通道的选择性不够强,因而重金属能通过钙离子通道进入植物体内[7]。LaCl3是公认的Ca 离子竞争性抑制剂,能抑制Ca 离子进入细胞原生质体。本实验与对照组相比,LaCl3处理组HC 和LC 的根部Ca 含量显著下降,Cr、Pb 含量亦显著降低(P <0.05)。添加钙离子通道抑制剂能显著抑制菜心品种根部对Cr、Pb 的吸收,说明Cr、Pb 也通过钙离子通道进入菜心根部。Cataldo 等[21]在研究大豆时也得到了类似的结果。薛艳等[21]在研究芦蒿时也发现,Cd、Pb 可能通过质膜上的钙离子通道进入细胞内。也有报道缺铁诱导增加了李氏禾对Cr 的吸收,由此推断李氏禾可通过铁转运体吸收Cr[22]。添加钾离子通道抑制剂能显著抑制小花南芥根对Pb 的吸收[23]。因此,Cr、Pb 可通过钙离子通道进入菜心根细胞,但不排除通过其他通道吸收Cr、Pb。本实验中由于Cr、Pb 同时通过钙离子通道进入根系,钙离子通道功能活跃的品种,吸收Cr、Pb 的机会就较多,可能导致高吸收;反之,钙离子通道功能不活跃的品种,吸收Cr、Pb 的机会就较少,可能导致低吸收。在本研究的水培实验中,与HC 相比,LC 菜心根系对Cr、Pb 累积量较少,表 6 显示LC 菜心根系吸收Ca 的数量也较少,表明相对低累积菜心品种钙离子通道功能可能不活跃,吸收Ca 和Cr、Pb 的能力亦较低。控制重金属从根系向可食部分转运的关键环节是从根系细胞原生质体向导管组织的跨质膜反转运。该过程通常不经由钙离子通道完成,而是通过一些金属反转运蛋白进行,如HMA 家族中的某些反转运蛋白[24]。本研究中Cr 从根到可食部分转运系数在盆栽条件下HC 的特级粗条、油青四九显著高于LC的绿星、极品,而水培条件下却是相反,绿星高于特级粗条、油青四九,极品高于油青四九。水培条件下Ca、Fe、Cu、Na、Mg 等金属均以溶解态存在,而盆栽条件下溶解态金属较少。因此,两种培养条件下根系周围的溶解态金属阳离子与Cr、Pb 比例不同,导致品种吸收转运Cr 的数量不同,从而引起了品种间Cr 转运系数的差异变化。ZHANG et al 通过不同Cd 浓度处理的水培、盆栽试验发现苋菜Amaranthushybridus L.的Cd转运系数分别为0.64~1.50 和0.33~0.92[25]。同一培养条件下,外源添加重金属浓度的不同也引起了转运系数的差异。因此,认为不同培养条件下重金属与环境中阳离子的比例可能影响作物对重金属的吸收,从而改变其对重金属的转运能力。Pb 转运系数在盆栽条件下HC/LC 之间无显著差异,在水培条件下LC 却显著高于HC,说明Pb 从根到可食部分的转运不是导致LC 可食部分Pb 含量低的原因。HC/LC 之间Cr、Pb的转运系数变化趋势不一致,可能是Cr、Pb 在根细胞跨质膜反转运的机制不同,也可能是它们在导管组织长距离运输中的迁移能力不同导致,具体机制需进一步研究。
4 结论(1) 绿星和极品为Cr/Pb 相对低累积品种,提倡在Cr、Pb 轻度污染耕地种植相对低累积品种。
(2) 推测品种Cr/Pb 低累积的根际机制可能在于:根际水溶态Cr、Pb、DOC和H+较少,较少Cr、Pb借钙离子通道进入根细胞内,低累积品种对土壤Cr、Pb活化、吸收能力较弱,进而减少其可食部分对Cr、Pb累积。
| [1] | 吴燕明, 吕高明, 周航, 等. 湘南某矿区蔬菜中Pb、Cd污染状况及健康风险评估[J]. 生态学报 , 2014, 34 (8) : 2146–2154. WU Yan-ming, LÜ Gao-ming, ZHOU Hang, et al. Contamination status of Pb and Cd and health risk assessment on vegetables in a mining area in Southern Hunan[J]. Acta Ecologica Sinica , 2014, 34 (8) : 2146–2154. |
| [2] | YU H, WANG J. L, FANG W, et al[J]. Science of the Total Environment , 2006, 370 (2/3) : 302–309. |
| [3] | Grant C A, larke J M, Duguid S, et al. Selection and breeding of plant cultivars to minimize cadmium accumulation[J]. Science of the Total Environment , 2008, 390 (2/3) : 301–310. |
| [4] | 龚玉莲, 杨中艺. 蕹菜不同镉积累品种的根际土壤化学特征[J]. 应用生态学报 , 2014, 25 (8) : 2377–2384. GONG Yu-liang, YANG Zhong-yi. Chemical characteristics of the rhi zosphere soil of water spinach cultivars differing in Cd accumulation[J]. Chinese Journal of Applied Ecology , 2014, 25 (8) : 2377–2384. |
| [5] | Huang B F, Xin J L, Dai H W, et al. Identification of low-Cdcultivars of sweet potato (Ipomoea batatas(L[J]. Environmental Science and Pollution Research , 2015, 22 (15) : 11813–11821. |
| [6] | He B, Ling L, Zhang L, et al. Cultivar-specific differences in heavy metal(Cd, Cr, Cu, Pb, and Zn)concentrations in water spinach(ipomoea aquatic‘forsk’)grown on metal-contaminated soil[J]. Plant and Soil , 2015, 386 (1/2) : 251–262. |
| [7] | 郭晓方, 卫泽斌, 丘锦荣, 等. 玉米对重金属累积与转运的品种间差异[J]. 生态与农村环境学报 , 2010, 26 (4) : 367–371. GUO Xiao-fang, WEI Ze-bin, QIU Jin-rong, et al. Differences between corn cultivars in accumulation and translocation of heavy metals[J]. Journal of Ecology and Rural Environment , 2010, 26 (4) : 367–371. |
| [8] | Qiu Q, Wang Y T, Yang Z Y, et al. Responses of different Chinese flow ering cabbage(Brassica parachinensis L[J]. Journal of Food Agriculture & Environment , 2011, 39 (11) : 925–932. |
| [9] | 万敏, 周卫, 林葆. 不同镉积累类型小麦根际土壤低分子量有机酸与镉的生物积累[J]. 植物营养与肥料学报 , 2003, 9 (3) : 331–336. WAN Min, ZHOU Wei, LIN Bao. Low-molecular-weight organic acids in rhizosphere soil and their effection cadmium accumulation in two cultivars of wheat[J]. Plant Nutrition and Fertilizer Science , 2003, 9 (3) : 331–336. |
| [10] | Mei X Q, Li S S, Li Q S, et al. Sodium chloride salinity reduces Cd up take by edible amaranth(Amaranthusmangostanus L[J]. Ecotoxicology and Environmental Safety , 2014, 105 : 59–64. |
| [11] | 薛艳, 王超, 王沛芳, 等. 外源添加抑制剂对芦蒿吸收Cd和Pb的影响[J]. 农业环境科学学报 , 2009, 28 (11) : 2255–2258. XUE Yan, WANG Chao, WANG Pei-fang, et al. Effect of additive in hibitor on the Cd and Pb uptake of Artemisia seleirgensis[J]. Journal of Agro-Environment Science , 2009, 28 (11) : 2255–2258. |
| [12] | 刘维涛, 周启星, 孙约兵, 等. 大白菜对铅积累与转运的品种差异研究[J]. 中国环境科学 , 2009, 29 (1) : 63–67. LIU Wei-tao, ZHOU Qi-xing, SUN Yue-bin, et al. Variety difference of lead accumulation and translocation in Chinese cabbage(Brassica peckinensis L.)[J]. China Environmental Science , 2009, 29 (1) : 63–67. |
| [13] | 杨暹, 郭巨先, 刘玉涛. 华南特产蔬菜菜心的营养成分及营养评价[J]. 食品科技 , 2002 (9) : 74–76. YANG Xian, GUO Ju-xian, LIU Yu-tao. The nutritive composition and assessment of Brassica parachinensis[J]. Food Science and Technology , 2002 (9) : 74–76. |
| [14] | 鲍士旦. 土壤农化分析[M].三版. 北京: 中国环境科学出版社, 2005 : 25 -241. BAO Shi-dan. Soil agro-chemistrical analysis[M].The third edition. Beijing: China Environmental Science Press, 2005 : 25 -241. |
| [15] | 国家环境保护总局. HJ 332-2006, 食用农产品产地环境质量评价标准[S]. 北京:中国环境科学出版社, 2007. State Environmental Protection Administration. HJ 332-2006, Farm land environmental quality evaluation standards for edible agricultural products[S]. Beijing:China Environmental Science Press, 2007. |
| [16] | 中国环境监测总站. 中国土壤元素背景值[M]. 北京: 中国环境科学出版社, 1990 . China Environmental Monitoring Station. Natural background values of soil elements in China[M]. Beijing: China Environmental Science Press, 1990 . |
| [17] | 国家卫生和计划生育委员会. GB 2762-2012食品安全国家标准食品中污染物限量[S]. 北京:中国标准出版社, 2012. National Health and Family Planning Commission. GB 2762-2012 China National Food Safety Standard:Maximum limit of contaminants in food[S]. Beijing:Standards press of China, 2012. |
| [18] | 张黛静, 邵云, 柴宝玲, 等. 小麦植株对重金属铬吸收累积的研究[J]. 农业环境科学学报 , 2009, 28 (2) : 311–315. ZHANG Dai-jing, SHAO Yun, CHAI Bao-ling, et al. Uptake and ac cumulation of chromium in wheat[J]. Journal of Agro-Environment Science , 2009, 28 (2) : 311–315. |
| [19] | Chen Y X, Yu M G, Duan D C. Tolerance, accumulation, and detoxifi cation mechanism of copper in Elsholtzia splendens Sherameti I, Varma A, eds[J]. Detoxification of heavy metal Springer , 2011 : 317–344. |
| [20] | Yang Y Y, Jung J Y, Song W Y, et al. Identification of rice varieties with high tolerance or sensitivity to lead and characterization of the mechanism of tolerance[J]. Plant Physiology , 2000, 124 (3) : 1019–1026. |
| [21] | Cataldo D A, Garland T R, Wildung R E. Cadmium uptake kinetics in intact soybean plants[J]. Plant Physiology , 1983, 73 : 844–848. |
| [22] | Liu J, Duan C Q, Zhang X H, et al. Characteristics of chromium (Ⅲ) uptake in hyperaccumulator Leersia hex andra Swartz[J]. Environmental and Experimental Botany , 2011, 74 : 122–126. |
| [23] | 王吉秀, 太光聪, 祖艳群, 等. 小花南芥根(Arabisalpinal var. parvi flora Franch)对铅锌吸收的药理学研究[J]. 中国农学通报 , 2011, 27 (9) : 206–211. WANG Ji-xiu, TAI Guang-cong, ZU Yan-qun, et al. A Pharmacology study on the absorption of lead and zinc in roots of Arabis alpinal var[J]. Chinese Agricultural Science Bulletin , 2011, 27 (9) : 206–211. |
| [24] | Pedas P, Schjoerring J K, Husted S. Identification and characterization of zinc-starvation-induced ZIP transporters from barley roots[J]. Plant Ph ys iology Biochemistry , 2009, 47 (5) : 377–383. |
| [25] | Zhang X C, Zhang S R, Xu X X, et al. Tolerance and accumulation characteristics of cadmium in Amaranthushybridus L[J]. Journal of Hazardousmaterials , 2010, 180 (1) : 303–308. |