 2016, Vol. 35
2016, Vol. 35文章信息
- 周碧青, 陈成榕, 张黎明, 杨文浩, 毛艳玲, 邢世和
- ZHOU Bi-qing, CHEN Cheng-rong, ZHANG Li-ming, YANG Wen-hao, MAO Yan-ling, XING Shi-he
- 亚热带不同果园土壤可溶性有机氮季节动态及其影响因素
- Seasonal dynamics and impact factors of soil soluble organic nitrogen in different subtropical fruit plantations
- 农业环境科学学报, 2016, 35(9): 1735-1741
- Journal of Agro-Environment Science, 2016, 35(9): 1735-1741
- http://dx.doi.org/10.11654/jaes.2016-0170
文章历史
- 收稿日期: 2016-02-02






2. 土壤生态系统健康与调控福建省高校重点实验室, 福州 350002 ;
3. School of Environment, Griffith University, Queensland Brisbane, Qld 4111
2. University Key Lab of Soil Ecosystem Health and Regulation in Fujian, Fujian Agriculture and Forestry University, Fuzhou 350002, China ;
3. School of Environment, Griffith University, Brisbane, QLD 4111, Australia
可溶性有机氮(Soluble organic nitrogen,SON)是指土壤中可溶于水、盐溶液(如1 mol·L-1 KCl、0.5mol·L-1 K2SO4和10 mmol·L-1 CaCl2 等)或用电超滤法(EUF)提取的有机态氮[1]。国内外研究表明,SON 是林地、草地、茶园和农田生态系统土壤可溶性氮(Totalsoluble nitrogen,TSN)的主要组分,其所占比例甚至超过NH4+、NO3-等可溶性无机氮[2-10];SON 是北美森林流域和西班牙东南部以农业为主集水区土壤氮素流失的主要形态,流域河水中SON 分别占TSN 的90%和72%~97%,严重影响流域水质[11-13];连续施用高量猪粪3 年后亚热带地区酸性玉米地土壤SON 淋失量显著提高[14]。可见,区域土壤SON 含量、季节动态及其迁移可能对流域水环境安全产生不利影响,且影响效应甚至超过人类至今仍普遍关注的NH4+、NO3-等可溶性无机氮。因此,研究探讨不同陆地生态系统土壤SON 的动态变化、差异及其影响因素,对于深入揭示陆地生态系统土壤SON 的行为过程和环境效应、防控SON 流失所致的氮素面源污染等具有重要的理论和现实意义。然而,国内外至今对陆地生态系统土壤SON 季节动态及其影响因素研究报道较少[15-16],有关不同类型果园土壤SON 季节动态差异及其影响因素研究则更少见报道。为此,本研究以福建南亚热带地形、母质、土壤类型和种植年限均相同的两个不同类型相邻果园(龙眼园和枇杷园)为研究对象,设立定时定位观测小区,借助TOC-TN 分析、离子自动分析和气相色谱分析等技术,分析不同果园生态系统、不同季节上下层土壤SON 含量及其相关理化和生化性质,研究探讨亚热带不同类型果园土壤SON 的季节动态差异及其影响因素,旨在为深入揭示陆地生态系统土壤SON 的行为过程及环境效应提供科学依据。
1 研究区与研究方法 1.1 研究区概况研究区位于福建省莆田市农科所果树研究中心果园,属南亚热带海洋性季风气候,年均气温20℃,年均日照时数1947 h,年无霜期320~350 d,年均降水量1300 mm,其中以春季降水量最高,占年均降水量的42.6%;自然坡度15毅,坡向东南,已垦为宽度3 m的果园梯台;成土母质为花岗岩坡残积物,土壤类型为硅铝质砖红壤性红壤。供试果园为种植于1993 年的相邻龙眼园和枇杷园,两种果园每年均施用化肥3次,其中龙眼园化肥施用量为180 kg N·hm-2·a-1、160kg P2O5·hm-2·a-1和225 kg K2O·hm-2·a-1,分别于每年的3 月(花前肥,占年化肥施用量的50%)、6 月(幼果肥,占20%)和9 月(采果肥,占30%)施用;枇杷园化肥施用量为360 kg N·hm-2·a-1、150 kg P2O5·hm-2·a-1和270 kg K2O·hm-2·a-1,分别于每年的2 月(幼果肥,占20%)、5 月(采果肥,占45%)和9 月(花前肥,占35%)施用。此外,龙眼园和枇杷园还分别于每年12月和6月以基肥方式施用有机肥,在果树树冠滴水线下开设环状沟,分别施用鸡粪肥(N、P2O5和K2O 含量分别为16.3、15.4 g·kg-1和9 g·kg-1)18 000 kg·hm-2·a-1和13 500 kg·hm-2·a-1。
1.2 研究方法 1.2.1 观测小区布设与样品采集在供试龙眼园和枇杷园中选择海拔相同(80 m)的果园梯台布设定时定位观测小区,小区面积为15m×20 m。每一果园设3个重复,在每一观测小区以“S”型布点法在树冠滴水线内确定采样点,并用标桩标记。于2010 年10 月—2012年7 月期间的秋季(10月)、冬季(1 月)、春季(4 月)和夏季(7 月)的中旬(每月15 日)在各观测小区分别定位采集0~20 cm(上层)和20~40 cm(下层)土样10 个,采样时注意避开果树施肥穴,分别将各小区同一层次10 个土样充分混匀后以四分法取得混合样1 kg,将各小区混合样分成2 份,其中一份样品(400 g)置于车载冰箱4℃存放,另一份样品(600 g)常温下存放。在实验室将4℃存放样品通过2 mm 筛后置于4℃冰箱恒温保存,供SON、微生物区系、相关酶活性和自然含水量测定,另一份样品进行风干处理,过2、1 mm 和0.149 mm 筛后常温下保存,供土壤相关理化性质测定。
1.2.2 分析方法土壤SON 通过总可溶性氮(TSN)与可溶性无机氮(Soluble inorganic nitrogen,SIN,即铵态氮和硝态氮之和)差减法计算得到,其中TSN 采用2 mol·L-1 KCl(土/水比为1:10)提取、SHIMADZU TOC-VCPH/CPN分析仪测定,铵态氮和硝态氮采用LACHATQuickchem 离子自动分析仪测定。采用SHIMADZUTOC 分析仪测定提取液中的总可溶性碳和可溶性无机碳含量,通过差减法计算得到可溶性有机碳(SOC)含量[17]。
土壤含水量、有机质、pH 值和质地分别采用105益烘干法、重铬酸钾容量法、酸度计法(土:水=1:2.5)和吸管法测定[18]。土壤微生物区系采用磷脂脂肪酸谱图分析法测定,土壤磷脂脂肪酸提取参考Frostegard方法[19],采用美国Agilent6890N型气相色谱仪内标法-MIDI 公司Sherlock MIS4.5 系统鉴定微生物区系[20],土壤细菌和真菌生物量按特征脂肪酸占总脂肪酸的mol%表示。土壤蛋白酶和天门冬酰胺酶活性分别采用茚三酮比色法和扩散法测定[21]。在各定时定位观测小区埋设自动记录地温计观测0~20 cm 和20~40 cm的土壤温度,每日间隔4 h自动测量记录地温一次,分别统计供试龙眼园和枇杷园各月0~20 cm 和20~40 cm土壤平均温度。
1.2.3 土壤SON 季节动态影响因素灰色关联分析土壤生态系统是一个灰色系统,其SON 动态是诸多影响因素综合作用的结果,各影响因素与土壤SON 含量间的关系不仅是灰色的,且影响程度也各异。灰色关联分析是基于灰色系统的灰色过程,是对一个系统发展变化态势的定量描述和比较,通过计算系统内各因子之间的关联度系数,可以揭示系统构成因素之间的关联性及其主次关系[22],故本研究采用灰色关联分析模型探讨果园土壤SON 季节动态的影响因素及其影响程度。由于供试两果园土壤相邻且地形、母质、土壤类型均相同,其土壤砂粒、粉粒、黏粒含量差异不大且不因季节而变化,故它们对果园土壤SON 季节动态影响不大,则可能影响果园土壤SON季节动态的因素包括有机质含量、细菌和真菌生物量、蛋白酶和天门冬酰胺酶活性以及pH 值、温度和含水量。以不同季节果园土壤SON 含量为母序列(n=96),可能影响因素为子序列(n=96),借助DPS14.5 数据处理系统的灰色关联分析模块进行不同季节果园土壤性质与SON 含量的关联分析,采用均值化法进行原始数据变换,分辨系数取0.5[22]。
1.3 数据统计分析借助SPSS 16.0 和Excel 2003 软件进行数据统计分析,采用方差分析和LSD 多重比较进行差异显著性检验。
2 结果与分析 2.1 不同类型果园土壤SON含量及SON/TSN差异从不同类型果园相同土层的SON 含量比较来看(图 1、图 2),春、夏、秋、冬季中旬龙眼园土壤SON 含量均大于枇杷园。其中龙眼园上层(0~20 cm)土壤春、夏、秋、冬季中旬的SON 含量分别比枇杷园上层土壤的高4.12、2.65、2.34、5.83 mg·kg-1。下层(20~40cm)土壤SON含量分别比枇杷园的高213.4%、16.0%、23.0%和20.7%,两者间差异达显著或极显著水平。

|
| 图 1 果园0~20 cm 土壤SON 含量(n=6) Figure 1 Soil SON contents in 0~20 cm layer(n=6) 柱状图上方小写字母和大写字母表示龙眼园与枇杷园之间土壤 SON含量差异分别达1%和5%显著水平。下同 |

|
| 图 2 果园20~40 cm 土壤SON 含量(n=6) Figure 2 Soil SON contents in 20~40 cm layer(n=6) |
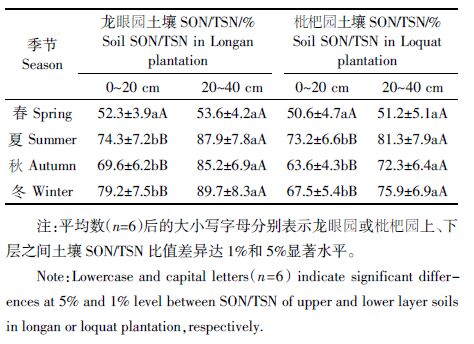
SON/TSN 反映了土壤SON 在可溶性氮组分中所占的比重大小。研究结果表明(表 1),供试龙眼园春、夏、秋、冬四季中旬上层(0~20 cm)土壤SON 含量占TSN 的52.3%耀79.2%,均值为68.8%,枇杷园上层(0~20 cm)土壤SON 含量占TSN 的50.6%耀73.2%,均值为63.7%;龙眼园春、夏、秋、冬四季中旬下层(20~40cm)土壤SON 含量占TSN 的53.6%耀89.7%,均值为79.1%,枇杷园下层(20~40 cm)土壤SON 含量占TSN的51.2%耀81.3%,均值为70.2%。可见,尽管不同季节、不同果园、不同深度土层SON/TSN 的比例各异,但土壤SON 占TSN 的比例均超过50%,SON/TSN 比例的年均值介于63.7%耀79.1%,表明SON 是供试龙眼园和枇杷园土壤可溶性氮的主要组成成分。从表 1还可看出,除春季中旬龙眼园和枇杷园上、下层土壤之间的SON/TSN 比值差异不显著外,夏、秋、冬季中旬龙眼园和枇杷园下层土壤的SON/TSN 比值均极显著高于上层土壤,说明SON 在两种果园下层土壤可溶性氮组分中所占比例更高,表明供试果园土壤SON存在向下迁移现象。
|
从不同类型果园土壤SON 含量季节动态分析可以看出(图 1、图 2),两种供试果园土壤SON 的季节变化均表现为夏、冬季高于春、秋季。其中龙眼园冬季中旬上、下层土壤SON 含量分别比春、秋和夏季中旬高15.60、6.98、1.77 mg·kg-1和8.55、2.52、0.09 mg·kg-1,枇杷园夏季中旬上、下层土壤SON 含量分别比春、秋和冬季中旬高15.30、4.90、1.41 mg·kg-1和11.24、2.74、0.45 mg·kg-1,夏、冬季中旬果园土壤SON 含量与春、秋季的差异达显著或极显著水平,秋季和春季间的差异也达极显著水平,而夏季与冬季间的差异则未达显著水平。可见,供试两种果园土壤SON 的季节动态均呈“N”型变化,龙眼园上、下层土壤SON 含量均以冬季最高,分别为26.65 mg·kg-1和15.82 mg·kg-1,枇杷园则以夏季最高,分别为22.23 mg·kg-1和13.56 mg·kg-1,且两种供试果园土壤SON 含量均以春季最低。
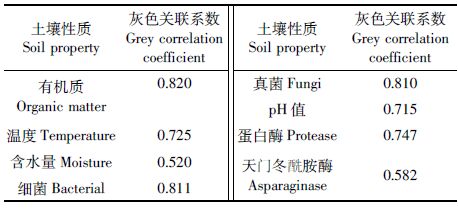
2.3 果园土壤SON 季节动态可能影响因素的灰色关联分析灰色系统理论通过关联度分析,理清系统中各因素间的主次关系,找出影响最大的因素,关联度是系统内两个因素之间关联性大小的量度,关联度愈大,系统内两个因素之间的关系就愈密切,相互影响程度也越大[22]。灰色关联分析结果可见(表 2),供试果园土壤SON 季节动态与可能影响因素的灰色关联系数大小呈以下次序:有机质含量跃细菌和真菌生物量跃蛋白酶活性跃温度跃pH值跃天门冬酰胺酶活性跃含水量。土壤有机质含量、细菌和真菌生物量是影响供试果园土壤SON 季节动态的关键因素,其灰色关联系数均≥0.810,主要通过作为SON 来源物和生产者而对果园土壤SON 季节动态产生显著影响;土壤蛋白酶活性、温度和pH 值是影响供试果园土壤SON 季节动态的重要因素,其灰色关联系数介于0.715~0.747,主要通过作为SON 形成的催化剂以及制约SON 生产者(细菌和真菌)活动而较显著影响果园土壤SON 季节动态变化;而土壤天门冬酰胺酶活性和含水量对果园土壤SON 季节动态影响程度相对较小,其灰色关联系数≤0.582,主要通过作为SON形成的催化剂、影响土壤通气性以及SON 淋失等影响果园土壤SON 季节动态变化。
|
土壤生态系统SON 含量受气候、土壤类型、覆被类型、管理措施、土壤理化和生化性质以及环境条件等诸多生物和非生物因素影响[1-6]。已有研究表明,中亚热带阔叶林土壤2 mol·L-1 KCl 提取的SON 含量显著高于针阔混交林和针叶林土壤,阔叶林、针阔混交林及针叶林0~20 cm 和20~40 cm 土壤2 mol·L-1 KCl提取SON 含量分别占TSN 的82.0%、77.1%、77.7%和74.4%、75.1%、78.5%[2];中亚热带乌龙茶园土壤2mol·L-1 KCl 提取SON 含量显著高于绿茶园,乌龙茶、绿茶园0~15 cm和15~30 cm 土壤2 mol·L-1 KCl 提取SON 含量分别占TSN 的80.4%、77.3%和63.8%、64.6%[3]。本研究结果表明,南亚热带龙眼园0~20 cm和20~40 cm 土壤2 mol·L-1 KCl 提取SON 含量显著或极显著高于枇杷园土壤;供试龙眼园春、夏、秋、冬季中旬0~20 cm 和20~40 cm 土壤SON 分别占TSN的52.3%~79.2%和53.6%~89.7%;枇杷园上、下层土壤SON 分别占TSN 的50.6%~73.2% 和51.2% ~81.3%。由此可见,SON 也是供试南亚热带果园土壤可溶性氮的主要组成成分,其含量因果园生态系统类型不同而差异明显,这与其他生态系统土壤SON 研究结果相一致[2-3, 5-8]。究其原因主要是由于不同类型果园土壤SON 来源物数量和土壤生化性质差异所致,调查发现供试龙眼园土壤年有机肥输入量比枇杷园高33.3%,年复一年的有机肥大量施用,导致龙眼园土壤有机质含量、细菌及真菌生物量和蛋白酶活性均值分别比枇杷园土壤高12.5%~25.9%、33.3% ~34.8%、13.5%~14.9%和7.98%~22.7%;灰色关联分析表明土壤有机质含量、细菌和真菌生物量是影响果园土壤SON 的关键因素,蛋白酶活性是影响果园土壤SON 的重要因素之一,而已有研究表明土壤SON 含量与有机质含量、细菌及真菌生物量和蛋白酶活性均呈极显著正相关[2-3],致使供试龙眼园土壤SON 含量显著高于枇杷园。本研究结果还表明,除春季中旬龙眼园和枇杷园上、下层土壤之间SON/TSN 比值差异因该季节是研究区雨季、土壤SON 淋失作用较强而不显著外,夏、秋、冬季中旬龙眼园和枇杷园下层土壤SON/TSN 比值均极显著高于上层土壤,说明SON 在两种果园下层土壤可溶性氮组分中所占比例更高,这是由于SON 具有较高的溶解性和较强的运动性[23],致使SON 易于向下迁移所致。国外研究表明,土壤SON 是北美森林流域和西班牙东南部以农业为主集水区土壤氮素流失的主要形态,并对流域水质产生显著影响[11-13],而亚热带地区属于多雨区,故亚热带陆地生态系统中占优势的土壤SON 迁移及其对流域地表水富营养化影响的研究应引起高度重视。
土壤SON 含量动态变化表现为土壤中复杂有机氮被微生物分解后释放出SON,产生的SON 被土壤吸附或进一步被微生物降解为无机氮,以及降解生成和人为施用的部分无机氮又被微生物同化为SON 等过程综合作用的结果。国内外至今有关生态系统土壤SON 季节动态变化的研究仍较少,且集中于林地土壤,研究结果也各异。Stevens等[24]研究酸性林地土壤SON季节变化时发现SON浓度随温度升高而增加,且土壤SON 季节动态表现为夏季高于冬季;Michalzik等[25]在调查挪威、德国温带森林土壤溶液中SON 含量时发现,SON 浓度主要表现为夏季较冬季高;但Huang 等[26]研究结果则相反,发现夏季因气温高,有机质矿化速率较高,土壤SON 浓度则相对较低。本研究结果表明,供试两种果园土壤SON 季节动态均呈夏、冬季高于春、秋季的“N”型变化规律,其中龙眼园上、下层土壤SON 含量均以冬季最高,枇杷园则以夏季最高,且两种果园土壤SON 含量均以春季最低。供试果园土壤SON 季节动态呈现上述变化规律的原因主要与其作为SON 动态变化关键影响因素的SON来源物数量(有机肥、枯枝落叶输入)和主要微生物区系数量(细菌和真菌生物量)以及作为SON 动态变化重要影响因素的土壤环境条件(温度)季节变化密切相关。据调查,供试龙眼园每年在采收结束后(12月)、枇杷园在每年采果后及夏梢抽生期(6 月)均较大量施用有机肥,且两种果树在冬季也均有枯枝落叶回归土壤而形成枯枝落叶层,故夏、冬季分别是枇杷园和龙眼园土壤有机物质输入量最大的季节。此外,据土壤温度观测数据统计,研究区7 月上层(0~20cm)和下层(20~40 cm)平均土温分别为31.3℃和29.5℃,1 月平均土温分别为14.2℃和15.1℃,多数土壤微生物活动的最适温度为15~45℃[27],表明研究区夏、冬季土壤热量状况(尤其是夏季)有利于微生物活动。因此,夏季枇杷园及冬季龙眼园土壤因施用有机肥所致的较大量有机物质输入和适宜土壤温度所致的微生物较旺盛活动,致使土壤中较丰富的复杂有机氮被大量转化为SON,而产生的SON一方面被土壤颗粒吸附,另一方面被微生物进一步矿化释放出NH4+和NO3-,但生成的部分NH4+和NO3-又可能被微生物同化固定而以氨基酸和其他有机氮形式进入土壤SON库[28],这些过程综合作用的结果导致枇杷园夏季、龙眼园冬季土壤SON含量较高。Michalzik等[15]研究温带森林生态系统穿透雨、枯枝落叶层渗滤液和土壤中SON 浓度后发现,枯枝落叶层渗滤液的SON 浓度最高,森林土壤凋落物输入量能影响不同季节土壤SON浓度,它们之间的相关性达显著水平;周碧青等[9-10]研究表明亚热带茶园土壤SON 含量与茶树凋落物数量和性质密切相关,故供试枇杷园和龙眼园冬季产生的枯枝落叶层渗滤液SON 的输入,也是导致冬季两种果园土壤SON 含量较高的原因。春季是研究区梅雨季节,据气象站观测数据,春季平均降水量高达564.2mm,占年均降水量的42.6%,是研究区年降水量最高的季节,其中4 月平均降水量达143.7 mm,占春季降水量的25.5%,致使龙眼园和枇杷园土壤含水量较高(土壤自然含水量分别高达20.21%和19.77%),土壤通气性相对较差,微生物活动也较弱,土壤SON 不仅形成数量少,且由于SON溶解性和移动性较强致使春季果园土壤SON淋失量大,从而导致春季两种果园土壤SON含量均最低。此外,国外研究还表明,土壤SON含量与根系代谢产物和分泌物密切相关[29-30],故供试果园土壤SON季节动态可能还与龙眼和枇杷根系分泌物季节变化有关,这有待于进一步深入研究。
4 结论SON 是供试南亚热带龙眼园和枇杷园土壤可溶性氮的主要成分,且春、夏、秋、冬季龙眼园土壤SON含量均显著高于枇杷园。龙眼园和枇杷园土壤SON季节动态均呈夏、冬季高于春、秋季的变化规律,其中龙眼园和枇杷园土壤SON含量分别以冬季和夏季最高,但均以春季最低。土壤有机质含量、细菌和真菌生物量变化是影响果园土壤SON 季节动态的关键因素,土壤蛋白酶活性、温度和pH值变化是影响果园土壤SON季节动态的重要因素。因此,供试果园土壤SON季节动态规律及其差异主要与SON 来源物数量(有机肥、枯枝落叶输入)、主要微生物区系数量(细菌和真菌生物量)以及土壤环境条件(温度)的季节变化密切相关。
| [1] | Murphy D V, Macdonald A J, Stockdale E A, et al. Soluble organic nitrogen in agricultural soils[J]. Biology and Fertility of Soils , 2000, 30 (5/6) : 374–387. |
| [2] | Xing S H, Chen C R, Zhou B Q, et al. Soil soluble organic nitrogen and active microbial characteristics under adjacent coniferous and broadleaf plantation forests[J]. Journal of Soils and Sediments , 2010, 10 (4) : 748–757. DOI:10.1007/s11368-009-0159-9 |
| [3] | Xing S H, Chen C R, Zhang H, et al. Genotype and slope position control on the availability of soil soluble organic nitrogen in tea plantations[J]. Biogeochemistry , 2011, 103 (1) : 245–261. |
| [4] | Zhou X Q, Chen C R, Wu H W, et al. Dynamics of soil extractable carbon and nitrogen under different cover crop residues[J]. Journal of Soils and Sediments , 2012, 12 (6) : 844–853. DOI:10.1007/s11368-012-0515-z |
| [5] | Zhong Z, Makeschin F. Soluble organic nitrogen in temperate forest soils[J]. Soil Biology and Biochemistry , 2003, 35 (2) : 333–338. DOI:10.1016/S0038-0717(02)00252-3 |
| [6] | 杨绒, 严德冀, 周建斌, 等. 黄土区不同类型土壤可溶性有机氮的含量及特性[J]. 生态学报 , 2007, 27 (4) : 1397–1403. YANG Rong, YAN De-yi, ZHOU Jian-bin, et al. Soluble organic nitrogen(SON) in different soils on the Loess Plateau of China[J]. Acta Ecologica Sinica , 2007, 27 (4) : 1397–1403. |
| [7] | Yang K, Zhu J J, Yan Q L, et al. Soil enzyme activities as potential indicators of soluble organic nitrogen pools in forest ecosystems of Northeast China[J]. Annals of Forest Science , 2012, 69 (7) : 795–803. DOI:10.1007/s13595-012-0198-z |
| [8] | Jones D L, Shannon D, Murphy D V, et al. Role of dissolved organic nitrogen(DON) in soil N cycling in grassland soils[J]. Soil Biology and Biochemistry , 2004, 36 (5) : 749–756. DOI:10.1016/j.soilbio.2004.01.003 |
| [9] | 周碧青, 张黎明, 毛艳玲, 等. 不同提取方法下茶园土壤可溶性有机氮的差异[J]. 福建农林大学学报 , 2013, 42 (3) : 317–322. ZHOU Bi-qing, ZHANG Li-ming, MAO Yan-ling, et al. Difference of soil soluble organic nitrogen concentration in tea plantation by various extractions[J]. Journal of Fujian Agriculture and Forestry University , 2013, 42 (3) : 317–322. |
| [10] | 周碧青, 陈成榕, 张黎明, 等. 茶树品种对亚热带茶园土壤可溶性有机氮组成的影响[J]. 农业环境科学学报 , 2015, 34 (6) : 1158–1165. ZHOU Bi-qing, CHEN Cheng-rong, ZHANG Li-ming, et al. Effects of tea genotype on soil soluble organic nitrogen fractions in subtropical tea plantations[J]. Journal of Agro-Environment Science , 2015, 34 (6) : 1158–1165. |
| [11] | Perakis S S, Hedin L O. Nitrogen loss from unpolluted South American forests mainly via dissolved organic compounds[J]. Nature , 2002, 415 : 416–419. DOI:10.1038/415416a |
| [12] | Perakis S S, Hedin L O. Fluxes and fates of nitrogen in soil of an unpolluted old-growth temperate forest, Southern Chile[J]. Ecology , 2001, 82 (8) : 2245–2260. DOI:10.1890/0012-9658(2001)082[2245:FAFONI]2.0.CO;2 |
| [13] | Miguel L H, Kevin H, Rosario J E. Distribution of dissolved inorganic and organic nitrogen in river water and ground water in an agriculturally-dominated catchment, South-East Spain[J]. Water, Air, and Soil Pollution , 2009, 198 (1) : 335–346. |
| [14] | Long G Q, Jiang Y J, Sun B. Seasonal and inter-annual variation of leaching of dissolved organic carbon and nitrogen under long-term manure application in an acidic clay soil in subtropical China[J]. Soil and Tillage Research , 2015, 146 : 270–278. DOI:10.1016/j.still.2014.09.020 |
| [15] | Michalzik B, Matzner E. Dynamics of dissolved organic nitrogen and carbon in a central European Norway spruce ecosystem[J]. European Journal of Soil Science , 1999, 50 (4) : 579–590. DOI:10.1046/j.1365-2389.1999.00267.x |
| [16] | Wang X L, Ye J, Gonzalez P, et al. The impact of organic farming on the soluble organic nitrogen pool in horticultural soil under open field and greenhouse conditions:A case study[J]. Soil Science and Plant Nutrition , 2013, 59 (2) : 237–247. DOI:10.1080/00380768.2013.770722 |
| [17] | Chen C R, Xu Z H, Keay P. Total soluble nitrogen in forest soils as determined by persulfate oxidation and by high temperature catalytic oxidation[J]. Australian Journal of Soil Research , 2005, 43 (4) : 515–523. DOI:10.1071/SR04132 |
| [18] | 鲁如坤. 土壤农业化学分析[M]. 北京: 中国科学技术出版社, 2000 : 421 -425. LU Ru-kun. Soil agro-chemistry analysis[M]. Beijing: Chinese Science and Technology Press, 2000 : 421 -425. |
| [19] | Frostegard A, Tunlid A, Bååth E. Microbial biomass measured as total lipid phosphate in soils of different organic content[J]. Journal of Microbiological Methods , 1991, 14 (3) : 151–163. DOI:10.1016/0167-7012(91)90018-L |
| [20] | 周碧青, 陈成榕, 张辉, 等. 不同覆被类型林地土壤微生物区系差异性研究[J]. 福建农林大学学报 , 2009, 38 (5) : 542–547. ZHOU Bi-qing, CHEN Cheng-rong, ZHANG Hui, et al. Micro-flora difference in forestry soils with various vegetation types[J]. Journal of Fujian Agriculture and Forestry University , 2009, 38 (5) : 542–547. |
| [21] | 关松荫, 张德生, 张志明. 土壤酶及其研究法[M]. 北京: 中国农业出版社, 1986 : 274 -319. GUAN Song-yin, ZHANG De-sheng, ZHANG Zhi-ming. Soil enzyme and research methodology[M]. Beijing: Chinese Agriculture Press, 1986 : 274 -319. |
| [22] | 唐启义. DPS数据处理系统(第三卷)[M]. 北京: 科学出版社, 2013 : 1261 -1270. TANG Qi-yi. DPS data processing system(Volume 3)[M]. Beijing: Science Press, 2013 : 1261 -1270. |
| [23] | 周建斌, 陈竹君, 郑险峰. 土壤可溶性有机氮及其在氮素供应及转化中的作用[J]. 土壤通报 , 2005, 36 (2) : 244–248. ZHOU Jian-bin, CHEN Zhu-jun, ZHENG Xian-feng. Soluble organic nitrogen in soil and its roles in the supply and transformation of N[J]. Chinese Journal of Soil Science , 2005, 36 (2) : 244–248. |
| [24] | Stevens P A, Wannop C P. Dissolved organic nitrogen and nitrate in an acid forest soil[J]. Plant Soil , 1987, 102 (1) : 137–139. DOI:10.1007/BF02370915 |
| [25] | Michalzik B, Kalbitz K, Park J H, et al. Fluxes and concentrations of dissolved organic carbon and nitrogen:A synthesis for temperate forests[J]. Biogeochemistry , 2001, 52 (2) : 173–205. DOI:10.1023/A:1006441620810 |
| [26] | Huang W Z, Schoenau J J. Fluxes of water-soluble nitrogen and phosphorus in the forest floor and surface mineral soil of a boreal aspen stand[J]. Geoderma , 1998, 81 (3/4) : 251–264. |
| [27] | 陈华癸. 土壤微生物学[M]. 上海: 上海科学技术出版社, 1979 : 53 -57. CHEN Hua-kui. Soil microbiology[M]. Shanghai: Shanghai Science and Technology Press, 1979 : 53 -57. |
| [28] | Chen C R, Xu Z H. Analysis and behavior of soluble organic nitrogen in forest soils[J]. Journal of Soils and Sediments , 2008, 8 (6) : 363–378. DOI:10.1007/s11368-008-0044-y |
| [29] | Neff J C, Chapin Ⅲ F S, Vitousek P M. Breaks in the cycle:Dissolved organic nitrogen in terrestrial ecosystems[J]. Frontiers in Ecology and the Environment , 2003, 1 (4) : 205–211. DOI:10.1890/1540-9295(2003)001[0205:BITCDO]2.0.CO;2 |
| [30] | Molina J A E, Clapp C E, Allmaras R R., et al. Simulation of nitrogen rhizodeposition and assimilation back into corn(Zea mays L.) roots[J]. Soil Biology and Biochemistry , 2005, 37 (1) : 93–100. DOI:10.1016/j.soilbio.2004.06.009 |