 2017, Vol. 36
2017, Vol. 36文章信息
- 许黎明, 成春燕, 卢汉浪, 莫祺晖, 韦星明
- XU Li-ming, CHENG Chun-yan, LU Han-lang, MO Qi-hui, WEI Xing-ming
- 木薯渣堆肥的生物酶活性与有机组分降解的关系
- Relationship between bioenzyme activities and degradation of organic substances during cassava dregs composting process
- 农业环境科学学报, 2017, 36(1): 198-204
- Journal of Agro-Environment Science, 2017, 36(1): 198-204
- http://dx.doi.org/10.11654/jaes.2016-0908
文章历史
- 收稿日期: 2016-07-11




木薯是广西的重要经济作物,木薯渣是木薯加工的主要固体废弃物,其中粗蛋白和脂肪含量低,木质粗纤维含量较高,主要成分是纤维素、半纤维素和木质素,富含氮、磷、钾、碳、钙、镁、硫、锌、锰、铜、铁和钠等矿物质[1]。木薯渣作为原料在工业上可用于制取乙醇,生物酶和有机酸等,在农业上可用于食用菌栽培和动物饲料,另外,木薯渣还可以发酵生产甲烷等[2]。实际上由于木薯渣在上述应用中的消耗有限,若没有得到及时有效的处理而大量囤积,将会对环境造成污染,给人和动物的健康埋下隐患[3-4]。高温好氧堆肥是资源化处理利用有机固体废弃物的有效途径,经济和技术的可行性高[5],而且生物有机肥已经被广泛证明具有减少或代替部分化肥、农药的使用[6-7],增强作物抗逆性[8],改善作物品质[9],改良土壤等优势[10]。因此,通过堆肥方式快速发酵腐熟木薯渣,可以将大量木薯渣制成无污染且富含有益微生物和矿质元素的生物有机肥,建立有机循环产业链,有效缓解环境压力。
木薯渣堆肥发酵腐熟是由微生物主导的温度变化过程,前期以中温型微生物菌群为主,中后期则高温型微生物菌群占优势[11],微生物分泌的酶是其中主要因素,因而对堆肥过程中微生物及其酶活性变化进行研究,有利于了解堆肥的生化过程和腐解机制[12-13]。农业行业标准《NY 609—2002 有机物料腐熟剂》也将纤维素酶、淀粉酶和蛋白酶的活性作为有机物料腐熟菌剂的产品技术指标。目前,国内对木薯渣发酵过程中生物酶的研究多集中在酶活含量的测定上[13-16],而对于发酵过程中生物酶活性和木薯渣的理化因子间关系的研究很少。本文通过对木薯渣接种复合微生物菌剂的堆肥化过程中上述3种酶的酶活性动态变化与堆肥理化因子变化之间的相关性进行研究,以期掌握木薯渣堆肥中主要生物酶活性变化趋势,建立堆肥生物酶活性变化与堆肥有机组分的降解过程、堆肥腐熟进程间的联系,发掘堆肥发酵腐熟过程的生物学指标,为木薯渣堆肥生产的工艺控制提供指导。
1 材料与方法 1.1 试验材料木薯渣堆肥混合料以木薯渣为主要原料,添加适量豆粕调节碳氮比(C/N)至30 左右,麦麸作为疏松剂,调节含水率为65%,得到木薯渣发酵混合料。木薯渣取自南宁市明阳淀粉厂,其相关成分质量分数为:纤维素32.47%,半纤维素11.03%,木质素8.33%,淀粉43.46%,粗蛋白2.83%。豆粕为饲料级,购于市场,粗蛋白成分质量分数为43.21%。
接种所用的外源微生物菌剂为本实验室筛选的复合微生物菌剂,主要包含枯草芽孢杆菌(Bacillus subtilis)、酿酒酵母(Saccharomyces cerevisiae)、嗜热侧孢霉(Sporotrichum thermophile)和放线菌(Actinomycetes)等,有效活菌数不低于5×108 CFU·g-1。
1.2 试验设计实验设2 个处理,不接种微生物菌剂(处理1);堆肥前期按液料质量比8译喷施接种复合微生物菌剂(处理2)。堆肥过程中进行人工翻堆。
1.3 样品采集在堆肥的第0、3、5、7、10、15、20、25、30、35、40、50 d取样,每次分别从堆体的上、中、下层分散取样,样品混合均匀后,部分低温保存,部分用于相关指标的测定。
1.4 测定内容与方法温度用SATOSK-110TRH-II型数显温度计测定。NH4+-N 用靛酚蓝比色法测定,NO3--N 用紫外分光光度法测定。半纤维素、纤维素、木质素用ANKOMA2000i型纤维素分析仪进行测定。纤维素酶酶活采用DNS比色法进行测定,蛋白酶酶活采用福林法进行测定,淀粉酶酶活采用I2-KI 分光光度法进行测定,具体测定方法参见《NY 609—2002 有机物料腐熟剂》和《QB/T1803—1993工业酶制剂通用实验方法》。
1.5 数据处理采用Microsoft Office Excel 和SPSS 软件进行数据处理和分析。
2 结果与分析 2.1 堆肥过程的温度变化木薯渣堆肥温度变化见图 1。堆肥过程可分为升温期、高温期、降温期和低温稳定期4个阶段。处理2在第4 d堆体温度升至62℃,进入高温分解阶段,第8d达到最大值71℃;高温期持续约15 d,在第25 d堆体温度逐渐下降,到第35 d温度接近室温,木薯渣堆肥进入腐熟阶段。处理1的堆体温度在第7 d升至60℃,第13 d达到70℃,第20 d开始下降,此后温度继续下降,到第35 d接近室温。

|
| 图 1 木薯渣堆肥过程中温度变化 Figure 1 Change of the temperature during cassava dregs composting |
对比木薯渣堆肥方式的不同处理,处理2 比处理1升温快,进入高温期早,且高温期持续时间长。但无论是处理2 还是处理1,其55℃以上堆体温度的持续时间均达到15 d以上,可以有效杀灭致病菌而达到堆肥无害化目的。
2.2 堆肥水解酶活性和理化因子间的关系 2.2.1 堆肥纤维素酶活性变化和纤维素、半纤维素、木质素含量变化纤维素是堆肥微生物的能源物质和碳源,而纤维素酶则是参与纤维素水解的关键酶。如图 2A 所示,处理2 在0~10 d 期间,纤维素酶活力逐渐升高,当堆肥温度升至60℃以上时,纤维素酶维持在较高的酶活水平,在第25 d 后,纤维素酶活力逐渐下降。处理1在堆肥初期酶活力略有下降,在第7 d后酶活上升,在第30 d达到峰值之后酶活逐渐下降,并趋于平缓,直至堆肥结束。

|
| 图 2 木薯渣堆肥过程中纤维素酶活性以及纤维素、半纤维素和木质素的变化 Figure 2 Change of cellulase activity,cellulose,hemicellulose and lignin during cassava dregs composting |
堆肥腐熟的关键是木质纤维素这类难分解物质的快速分解[17]。
堆肥纤维素含量变化见图 2B。处理1 和处理2的纤维素含量在堆肥过程中均呈下降趋势。堆肥初始,堆体中纤维素含量为(31.37±0.46)%。处理2在第15 d其纤维素含量为(14.06±0.37)%,堆肥结束时堆体中纤维素含量为(11.79±0.39)%,处理2 纤维素的降解主要集中于堆肥的前15 d,而在堆肥第35 d 后,纤维素的分解缓慢;处理1 在第25 d堆体中纤维素含量为(18.79±0.41)%,堆肥结束时纤维素含量为(17.53±0.48)%,处理1 纤维素的分解主要集中于堆肥的前25 d,而在堆肥第25 d 后,纤维素的分解缓慢。以上数据表明,木薯渣堆肥化过程中,纤维素的降解集中于堆肥高温期,纤维素降解相对较快,之后降解速度逐渐趋于平稳。
堆肥半纤维素含量变化见图 2C。木薯渣堆肥半纤维素初始含量为(10.66±0.22)%,处理2 在第15 d其半纤维素含量为(3.35±0.33)%,堆肥结束时为(1.85±00.32)%;处理1 在第25 d其半纤维素含量为(5.09±0.42)%,堆肥结束时为(2.01±0.26)%。半纤维素相对于纤维素较容易降解,其分解也集中于堆肥过程中的高温期,在堆肥后期,半纤维素含量在堆肥中比较稳定。
堆肥木质素含量变化见图 2D。木质素一直是自然界中极难分解的物质之一,本研究中无论是处理1还是处理2,木质素的降解都非常低。
2.2.2 堆肥淀粉酶活力变化和淀粉含量变化木薯渣堆肥淀粉酶活力变化如图 3A。处理2 在第10 d时淀粉酶活力达到最大值,在第10~30 d,淀粉酶活力逐渐下降;处理1 在第15 d淀粉酶活力达到最大值,之后酶活力逐渐下降。

|
| 图 3 木薯渣堆肥过程中淀粉酶活性和淀粉的变化 Figure 3 Change of amylase activity and starch during cassava dregs composting |
堆肥淀粉含量变化见图 3B。处理1 和处理2 的淀粉含量在堆肥过程中均呈下降趋势。堆肥初始,堆体中淀粉的含量较高,为(41.99±0.52)%。处理2在第15 d其淀粉含量为(8.55±0.49)%,堆肥结束时堆体中的淀粉含量为(3.2±0.53)%;处理1 在第25 d其淀粉含量为(10.25±0.52)%,堆肥结束时堆体中的淀粉含量为(5.71±0.45)%。以上数据表明,木薯渣堆肥化过程中,淀粉的降解集中于堆肥高温期,淀粉降解相对较快,之后降解速度逐渐趋于平稳。
2.2.3 堆肥蛋白酶活性变化和NO3--N、NH4+-N含量变化木薯渣堆肥蛋白酶活力变化如图 4A 所示。蛋白酶酶活性在堆肥初期达到最大值,随后快速降低,第10 d后趋于平稳,整体变化较为平缓。

|
| 图 4 木薯渣堆肥过程中蛋白酶活性、NO3--N 和NH4+-N 的变化 Figure 4 Change of protease activity,nitrate nitrogen(NO3--N)and ammonia nitrogen(NH4+-N)during cassava dregs composting |
木薯渣堆肥NH4+-N 和NO3- -N 的含量变化见图 4B 和图 4C。木薯渣堆肥NH4+-N 含量整体呈下降趋势,在堆肥第5~10 d时段内快速降低;NO3--N 含量整体呈上升趋势,在堆肥第5~10 d时段内快速增长。堆肥的氮素变化中NO3--N 和NH4+-N 是评价堆肥腐熟度的常用指标。在本研究中,堆肥结束时NO3--N 为2.89 g·kg-1,NH4+ -N 的含量降至1.22 g·kg-1,NH4+ -N/NO3--N 的比值为0.42,与文献援引的堆肥腐熟评价标准NO3--N/NH4+-N 的比值≥2 或NH4+-N/NO3--N 的比值≤0.5相符[18]。
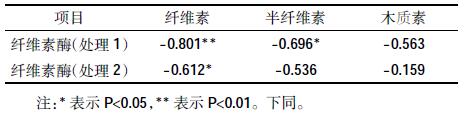
2.2.4 纤维素酶活性与纤维素、半纤维素和木质素的相关性分析由表 1可见,木薯渣堆肥处理1 的纤维素酶活性变化与纤维素含量为极显著负相关(r=-0.801),与半纤维素含量为显著负相关(r=-0.696),处理2 的纤维素酶活性变化与堆肥中的纤维素含量为显著负相关(r=-0.612),说明木薯渣堆肥纤维素酶活性变化与纤维素的降解显著相关。
 |
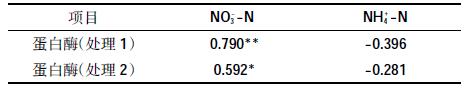
木薯渣堆肥处理1 的蛋白酶活性变化与NO3--N含量为极显著正相关(r=0.790),处理2 的蛋白酶活性变化与堆肥中的NO3- -N 含量为显著正相关(r=0.592),说明木薯渣堆肥蛋白酶活性变化与NO3--N的累积显著相关,参与堆肥氮素循环(表 2)。
 |
木薯渣堆肥处理1 的淀粉酶活性变化与淀粉含量为显著负相关(r=-0.584),处理2 的淀粉酶活性变化与淀粉含量显著负相关(r=-0.676),说明木薯渣堆肥淀粉酶活性变化与淀粉的降解显著相关(表 3)。

本研究结果表明,木薯渣堆肥的纤维素酶、淀粉酶和蛋白酶活性与堆肥物料特性相关,堆肥前期物料中较丰富的有机组分和适宜的温度促进了微生物的生长和产酶活力。经微生物的发酵作用,堆肥物料迅速分解,放热升温,第4 d就能迅速升温至62℃,纤维素酶、淀粉酶和蛋白酶的酶活性也随之升高,随着堆肥微生态改变,适温微生物富集繁殖,菌群生长代谢旺盛,消耗了大量纤维素、淀粉和蛋白质等有机组分,大量放热,堆肥维持高温阶段15 d 左右,纤维素酶和淀粉酶也随之维持一段时期的高酶活性状态,之后有机组分因大量消耗而匮乏,微生物生长代谢活动趋于平缓,温度逐渐降至气温,在30 d后进入低温腐熟期,完成堆肥化过程。这与侯宪文等[19]的研究一致。木薯渣堆肥中蛋白酶的酶活性前期变化短促,其原因是木薯渣自身蛋白质含量低,堆料中添加了有机氮源,在堆肥初期促进了产蛋白酶微生物的繁殖,蛋白酶活性快速上升,之后堆肥过程因大量含氮有机组分被消耗,蛋白酶活性也随之迅速降低。木薯渣堆肥2个处理间进行比较,处理2 的发酵周期比处理1 的周期短,并且堆肥纤维素、半纤维素的分解效率,以及NO3--N 的生成效率都有所提高,说明接种微生物菌剂堆肥发酵木薯渣可以达到比较好的效果。
木薯渣堆肥过程中纤维素酶、淀粉酶和蛋白酶的最高活性出现在堆肥的高温期,酶活变化基本上呈前期小幅缓慢调整,然后迅速上升,逐步缓慢下降,最后达到稳定的趋势,与堆肥过程中微生物生长分解代谢有机物质的特征有关[20-21]。堆肥中的酶活主要与物料中的微生物相关,来源于微生物的新陈代谢过程,木薯渣堆肥中有机物料发酵是在微生物菌群产生的各种酶的参与下进行的。
本研究中木薯渣堆肥淀粉酶酶活性与淀粉的含量呈显著负相关,说明木薯渣堆肥中淀粉酶主导了淀粉类物质的降解。
对木质纤维素的降解研究发现,纤维素酶和半纤维素酶同时破坏木质纤维素的不同结构有利于木质纤维素的酶解[22]。本研究中纤维素酶酶活性与纤维素的含量呈显著负相关,在木薯渣堆肥纤维素降解中起重要作用,而纤维素酶酶活性与半纤维素含量的关系在处理1 中呈显著负相关,处理2 则无显著相关性。分析其原因可能是处理2接种了外源微生物菌剂,整个堆肥发酵过程中的纤维素酶活性显著高于作为对照的处理1,而纤维素酶与半纤维素酶复合酶解木质纤维素是在一定范围内具有协同作用,其酶解率随着复合酶含量的增加而相应增加,一旦超出了范围,酶解率随着复合酶含量的持续增加其增加幅度下降[23]。由于木质素相比纤维素和半纤维素更难于分解,在本研究中木薯渣堆肥的纤维素酶活性与木质素含量之间无显著相关性。
堆肥氮素矿化是微生物驱动的生物化学过程,本研究中微生物蛋白酶参与堆肥中含氮有机组分的转化,其酶活性变化与堆肥NO3--N 含量呈显著正相关,但与NH4+-N 的含量无显著相关性。其原因可能是堆肥前期高温条件,以及强烈的蛋白酶作用和氨化作用将有机氮快速矿化分解,促进了NH3/NH4+-N 平衡向生成NH3方向移动,再加上频繁的人工翻堆通气散热,加速了堆体中NH3的流失,且翻堆后堆体会维持一定时间的低温状态,有利于硝化作用的进行,推动NH4+-N 向NO3--N 的转化。这些因素都造成了堆肥中NH4+-N 含量的下降和NO3--N 含量的累积。堆肥中后期含氮有机质因前期的大量消耗而匮乏,微生物蛋白酶活性下降,温度下降,硝化作用占主导,堆肥中的NH4+-N 进一步转化为NO2-和NO3- [19, 24-26]。另外,堆肥中后期温度下降,翻堆频次降低,堆体通风状况不畅,形成局部厌氧环境,反硝化作用促使NO3--N 生成N2O和N2,两方面原因造成了堆肥中后期NO3--N 总体累积放缓的趋势。
由此可见,本研究中堆肥材料和堆肥工艺影响了堆肥生物酶酶活性变化与堆肥有机组分降解。堆肥过程中的多种生物酶活性与C、N 等基础物质的代谢密切相关[13-14, 27],可在一定程度上反映堆肥腐熟度。本研究中堆肥纤维素酶、淀粉酶分别与纤维素、淀粉的发酵降解呈显著负相关,说明纤维素酶和淀粉酶参与堆肥碳素循环,分别主导了木薯渣堆肥中纤维素和淀粉的降解,故生物酶可以表征木薯渣堆肥中有机组分的分解状况[28-29]。另外,蛋白酶活性与氮素状况有极其重要的关系[30],堆肥中蛋白酶与NO3- -N 的含量呈显著正相关,说明蛋白酶参与了木薯渣堆肥氮素循环,表征堆肥NO3--N 的积累,而NO3--N 含量可以作为堆肥稳定性和腐熟度评价的指标。原因是在堆肥过程中,前期的氨化作用占优,NO3--N 含量较低,进入中后期硝化作用得到强化,部分NH4+-N 被转化为NO3--N 而使其含量迅速提高[19, 24]并趋于稳定,可以认为当堆肥中NO3--N 含量趋于稳定时即表明堆肥的状态已达稳定。因此,纤维素酶、淀粉酶和蛋白酶等生物酶的活性变化趋势可以表征木薯渣堆肥发酵腐熟的进程,在木薯渣堆肥中可以据此建立关键的生物酶活性指标产品品质监控节点,控制木薯渣堆肥的生产和评价堆肥的腐熟度。
4 结论本研究木薯渣堆肥中纤维素、淀粉和蛋白质等有机组分的降解主要集中在堆肥的高温期,其中接种微生物菌剂的木薯渣堆肥发酵处理方式效率较高;木薯渣堆肥过程中的生物酶活性与有机组分的降解呈显著或极显著相关关系,其中纤维素酶、淀粉酶和蛋白酶的活性变化趋势分别指示纤维素、淀粉的降解和NO3--N 的生成积累,说明生物酶活性可以共同表征木薯渣堆肥的发酵腐熟进程,可作为木薯渣堆肥发酵的生物活性质控指标。
| [1] | Barana A C, Cereda M P. Cassava wastewater(manipueira) treatment using a two-phase anaerobic biodigestor[J]. Food Science and Technology(Campinas), 2000, 20(2) : 183–186. |
| [2] | 李静, 涂佳才, 陈秀龙, 等. 木薯渣微生物降解及再利用研究进展[J]. 生态环境学报, 2010, 19(10) : 2506–2510. LI Jing, TU Jia-cai, CHEN Xiu-long, et al. Advances in reuse and microbiological degradation of cassava residue[J]. Ecology and Environmental Sciences, 2010, 19(10) : 2506–2510. |
| [3] | Ubalua A O. Cassava wastes:Treatment options and value addition alternatives[J]. African Journal of Biotechnology, 2007, 6(18) : 2065–2073. DOI:10.5897/AJB |
| [4] | Arotupin D J. Evaluation of microorganisms from cassava waste water for production of amylase and cellulose[J]. Research Journal of Microbiology, 2007, 2(5) : 475–480. DOI:10.3923/jm.2007.475.480 |
| [5] | 杨国义, 夏钟文, 李芳柏, 等. 不同填充料对猪粪堆肥腐熟过程的影响[J]. 土壤肥料, 2003(3) : 29–33. YANG Guo-yi, XIA Zhong-wen, LI Fang-bai, et al. Effect of different bulking agents on the maturity of pig manure composting[J]. Soils and Fertilizers, 2003(3) : 29–33. |
| [6] | El-Kramany M F, Bahr A A, Mohamed M F, et al. Utilization of bio-fertilizers in field crops production 16-groundnut yield, its components and seeds content as affected by partial replacement of chemical fertilizers by bio-organic fertilizers[J]. Journal of Applied Sciences Research, 2007, 3(1) : 25–29. |
| [7] | 袁英英, 李敏清, 胡伟, 等. 生物有机肥对番茄青枯病的防效及对土壤微生物的影响[J]. 农业环境科学学报, 2011, 30(7) : 1344–1350. YUAN Ying-ying, LI Min-qing, HU Wei, et al. Effect of biological organic fertilizer on tomato bacterial wilt and soil microorganism[J]. Journal of Agro-Environment Science, 2011, 30(7) : 1344–1350. |
| [8] | 凌宁, 王秋君, 杨兴明, 等. 根际施用微生物有机肥防治连作西瓜枯萎病研究[J]. 植物营养与肥料学报, 2009, 15(5) : 1136–1141. LING Ning, WANG Qiu-jun, YANG Xing-ming, et al. Control of Fusarium with watermelon by nursery application of bio-organic fertilizer[J]. Plant Nutrition and Fertilizer Science, 2009, 15(5) : 1136–1141. |
| [9] | 张建国, 聂俊华, 杜振宇. 复合生物有机肥在烤烟生产中的应用研究[J]. 植物营养与肥料学报, 2004, 10(4) : 424–428. ZHANG Jian-guo, NIE Jun-hua, DU Zhen-yu. Response of flue-cured tobacco to compound bio-organic fertilizer[J]. Plant Nutrition and Fertilizer Science, 2004, 10(4) : 424–428. |
| [10] | 王立刚, 李维炯, 邱建军, 等. 生物有机肥对作物生长, 土壤肥力及产量的效应研究[J]. 土壤肥料, 2004(5) : 12–16. WANG Li-gang, LI Wei-jiong, QIU Jian-jun, et al. Effect of biological organic fertilizer on crops growth, soil fertility and yield[J]. Soils and Fertilizers, 2004(5) : 12–16. |
| [11] | 吴圣进, 韦仕岩, 王灿琴, 等. 采用CFDA荧光染色技术监测木薯渣堆肥中的细菌动态[J]. 南方农业学报, 2012, 43(12) : 1976–1980. WU Sheng-jin, WEI Shi-yan, WANG Can-qin, et al. Bacteria detection in cassava residue compost by CFDA fluorescent staining technique[J]. Journal of Southern Agriculture, 2012, 43(12) : 1976–1980. |
| [12] | 戴芳, 曾光明, 牛承岗, 等. 堆肥化过程中生物酶活性的研究进展[J]. 中国生物工程杂志, 2005, 25(B04) : 148–151. DAI Fang, ZENG Guang-ming, NIU Cheng-gang, et al. Advance in the studies on bioenzyme activity during the composting[J]. China Biotechnology, 2005, 25(B04) : 148–151. |
| [13] | 谷洁, 李生秀, 秦清军, 等. 水解类酶活性在农业废弃物静态高温堆腐过程中的变化[J]. 中国农学通报, 2005, 21(5) : 32–35. GU Jie, LI Sheng-xiu, QIN Qing-jun, et al. Changes of some hydrolase activities during agricultural waste materials composting in high temperature and static state[J]. Chinese Agricultural Science Bulletin, 2005, 21(5) : 32–35. |
| [14] | 何江, 毛忠贵, 张庆华, 等. 高效木薯渣分解复合菌群RXS的构建及其发酵特性研究[J]. 环境科学, 2012, 33(3) : 1020–1027. HE Jiang, MAO Zhong-gui, ZHANG Qing-hua, et al. Construction of a microbial consortium RXS with high degradation ability for cassava residues and studies on its fermentative characteristics[J]. Environmental Science, 2012, 33(3) : 1020–1027. |
| [15] | 汤小朋, 赵华, 汤加勇, 等. 黑曲霉固态发酵改善木薯渣品质的研究[J]. 动物营养学报, 2014, 26(7) : 2026–2034. TANG Xiao-peng, ZHAO Hua, TANG Jia-yong, et al. Quality improvement of cassava residue by solid-state fermentation with Aspergillus niger[J]. Chinese Journal of Animal Nutrition, 2014, 26(7) : 2026–2034. |
| [16] | 肖马娜, 许曼, 林泽芬, 等. 木薯渣降解菌发酵条件及产酶特性研究[J]. 广东化工, 2015, 42(24) : 33–34. XIAO Ma-na, XU Man, LIN Ze-fen, et al. Research of enzyme-production characteristics and fermentation conditions of cassavadreg degrading fungi[J]. Guangdong Chemical Industry, 2015, 42(24) : 33–34. |
| [17] | 王伟东, 刘建斌, 牛俊玲, 等. 堆肥化过程中微生物群落的动态及接菌剂的应用效果[J]. 农业工程学报, 2006, 22(4) : 148–152. WANG Wei-dong, LIU Jian-bin, NIU Jun-ling, et al. Dynamics of microbial community and the effect of inoculum on the composting[J]. Transactions of the Chinese Society of Agricultural Engineering, 2006, 22(4) : 148–152. |
| [18] | 黄国锋, 钟流举, 张振钿, 等. 有机固体废弃物堆肥的物质变化及腐熟度评价[J]. 应用生态学报, 2003, 14(5) : 813–818. HUANG Guo-feng, ZHONG Liu-ju, ZHANG Zhen-tian, et al. Physicochemical changes and maturity evaluation of solid organic waste compost[J]. Chinese Journal of Applied Ecology, 2003, 14(5) : 813–818. |
| [19] | 侯宪文, 陈炫, 李勤奋, 等. 木薯渣堆肥过程中氮素转化及堆肥周期研究[J]. 中国农学通报, 2015, 31(26) : 145–149. HOU Xian-wen, CHEN Xuan, LI Qin-fen, et al. Study on nitrogen transformation and composting period in the process of cassava residue compost[J]. Chinese Agricultural Science Bulletin, 2015, 31(26) : 145–149. |
| [20] | Tiquia S M, Tam N F Y. Co-composting of spent pig litter and sludge with forced-aeration[J]. Bioresource Technology, 2000, 72(1) : 1–7. DOI:10.1016/S0960-8524(99)90092-5 |
| [21] | 张莹, 许修宏, 李洪涛. 污泥堆肥过程中生物酶活性与理化因子关系分析[J]. 农业环境科学学报, 2012, 31(11) : 2290–2295. ZHANG Ying, XU Xiu-hong, LI Hong-tao. Relationship between bioenzyme activities and physico-chemical properties during sewage sludge composting process[J]. Journal of Agro-Environment Science, 2012, 31(11) : 2290–2295. |
| [22] | Himmel M E, Ding S Y, Johnson D K, et al. Biomass recalcitrance:engineering plants and enzymes for biofuels production[J]. Science, 2007, 315(5813) : 804–807. DOI:10.1126/science.1137016 |
| [23] | 邵丽杰, 寇巍, 曹焱鑫, 等. 纤维素酶、半纤维素酶降解膨化玉米秸秆工艺优化[J]. 环境工程学报, 2014, 8(10) : 4373–4378. SHAO Li-jie, KOU Wei, CAO Yan-xin, et al. Optimization of degradation of expanded corn straw by cellulase and hemicellulose[J]. Chinese Journal of Environmental Engineering, 2014, 8(10) : 4373–4378. |
| [24] | 杨延梅, 刘鸿亮, 杨志峰, 等. 控制堆肥过程中氮素损失的途径和方法综述[J]. 北京师范大学学报:自然科学版, 2005, 41(2) : 213–216. YANG Yan-mei, LIU Hong-liang, YANG Zhi-feng, et al. Methods and techniques in the control of nitrogen loss during the composting:A review[J]. Journal of Beijing Normal University(Natural Science), 2005, 41(2) : 213–216. |
| [25] | 张晶, 林先贵, 尹睿. 参与土壤氮素循环的微生物功能基因多样性研究进展[J]. 中国生态农业学报, 2009, 17(5) : 1029–1034. ZHANG Jing, LIN Xian-gui, YIN Rui. Advances in functional gene diversity of microorganism in relation to soil nitrogen cycling[J]. Chinese Journal of Eco-Agriculture, 2009, 17(5) : 1029–1034. DOI:10.3724/SP.J.1011.2009.01029 |
| [26] | 贺纪正, 张丽梅. 土壤氮素转化的关键微生物过程及机制[J]. 微生物学通报, 2013, 40(1) : 98–108. HE Ji-zheng, ZHANG Li-mei. Key processes and microbial mechanisms of soil nitrogen transformation[J]. Microbiology China, 2013, 40(1) : 98–108. |
| [27] | 伍兵, 邓悟森, 谭周进, 等. 鸡粪堆肥的酶活特性研究[J]. 中国微生态学杂志, 2011, 23(11) : 972–975. WU Bing, DENG Wu-sen, TAN Zhou-jin, et al. Studies on enzyme characteristics during composting of chicken manure[J]. Chinese Journal of Microecology, 2011, 23(11) : 972–975. |
| [28] | Chefetz B, Kerem Z, Chen Y, et al. Isolation and partial characterization of laccase from a thermophilic composted municipal solid waste[J]. Soil Biology & Biochemistry, 1998, 30(8) : 1091–1098. |
| [29] | Dick R P. A review:long-term effects of agricultural systems on soil biochemical and microbial parameters[J]. Agriculture, Ecosystems & Environment, 1992, 40(1) : 25–36. |
| [30] | 徐惠风, 刘兴土. 长白山区沟谷沼泽乌拉苔草(Carexmeyeriana)湿地土壤酶活性与氮素、土壤微生物相关性研究[J]. 农业环境科学学报, 2009, 28(5) : 946–950. XU Hui-feng, LIU Xing-tu. Relationship between soil enzyme activity and nitrogen, soil microorganisms of Carexmeyeriana wetland in Changbaimountain valley[J]. Journal of Agro-Environment Science, 2009, 28(5) : 946–950. |