 2017, Vol. 36
2017, Vol. 36文章信息
- 穆海蓉, 邵在胜, 沈士博, 景立权, 王云霞, 王余龙, 杨连新
- MU Hai-rong, SHAO Zai-sheng, SHEN Shi-bo, JING Li-quan, WANG Yun-xia, WANG Yu-long, YANG Lian-xin
- 臭氧浓度增加对超级稻南粳9108稻穗不同部位籽粒氨基酸含量的影响
- Impacts of ozone stress on grain amino acids of super rice cultivar Nanjing 9108 differ with grain positions on a panicle
- 农业环境科学学报, 2017, 36(3): 420-427
- Journal of Agro-Environment Science, 2017, 36(3): 420-427
- http://dx.doi.org/10.11654/jaes.2016-1347
文章历史
- 收稿日期: 2016-10-24






2. 扬州大学环境科学与工程学院, 江苏 扬州 225009
2. College of Environmental Science and Engineering, Yangzhou University, Yangzhou 225009, China
在许多亚洲国家,近年来快速的经济发展使得空气污染物——氮氧化物和挥发性有机化合物排放增加,这些臭氧前体物在高温和强光照条件下通过光化学反应转化为臭氧,导致许多国家地表(近地层或对流层)臭氧浓度迅速上升,特别是人口密度大的发展中国家[1-2]。据预测,在未来的40年里,东南亚地区地表平均臭氧浓度将比现在高25 nL·L-1[2]。作为强氧化剂,目前地表臭氧浓度已超过敏感作物臭氧伤害阀值(40 nL·L-1)[3],对作物包括水稻的生长发育造成负面影响,未来这种影响可能更大[4-8]。
水稻作为人类最重要的食物来源,近年来国内外开展了大量有关臭氧浓度增加对水稻影响的研究,但多数集中在生长发育和产量形成方面,很少涉及稻米品质[5-7, 9]。地表臭氧浓度增高导致作物一系列生理过程发生变化,进而对植株物理或化学性状产生影响,收获器官的品质可能因此改变[7]。前期有限的研究表明,臭氧浓度增加对稻米品质的负面影响较大,影响程度与供试品种和气象条件有关[10-14]。稻米营养品质主要包括蛋白质含量及其氨基酸组成,是最为重要的品质性状之一。目前臭氧浓度增加对稻米营养品质的影响主要聚焦于蛋白质含量[10-11, 15-17],而对各种氨基酸包括重要的限制性氨基酸浓度的变化鲜有报道[17]。最新大田研究表明,臭氧浓度增加使不同密度水稻稻米氨基酸浓度均呈增加趋势[17],但这种趋势是否与籽粒在稻穗上的着生部位有关未见报道。水稻是穗状花序,同一穗内的颖花分化和开花存在先后,一般位于稻穗上部的强势粒开花早,灌浆启动快,灌浆时间短,而弱势粒则相反[18]。基于这种灌浆快慢的差异推测,臭氧浓度增加对弱势粒的影响可能大于强势粒,但这一假设尚未得到验证。
1996年中国实施超级稻育种计划以来,超级稻的选育取得重大进展,分别于2000、2004、2011、2014年实现第一至四期单季水稻产量目标[19-20],为我国水稻增产做出了重要贡献,但臭氧浓度增加对超级稻生长和产量的影响鲜有报道[21-22],稻米品质方面的研究则还是空白。本试验依托接近自然稻田环境的自然光气体熏蒸平台[23],以优质超级稻南粳9108为供试材料,用自然采光和群体土培方式培育水稻植株。试验设置对照(18 nL·L-1)和高浓度臭氧(100 nL·L-1)两个处理,成熟期籽粒按稻穗不同部位收获后测定糙米中的氨基酸,研究臭氧浓度增加对超级稻南粳9108氨基酸含量的影响及其与籽粒着生部位的关系,以期为高臭氧浓度环境下稻米营养品质的调控提供依据。
1 材料与方法 1.1 试验平台本试验依托一种新型自然光气体熏蒸平台[23],该平台位于江苏省扬州大学农学院网室试验田内(119.42°E,32.39°N)。试验土壤类型为清泥土,所在地年均降水量1000 mm左右,年均蒸发量在940 mm左右,年平均温度15 ℃,日照时间大于2100 h,年平均无霜期220 d。土壤理化性质:有机质24.3 g·kg-1,全氮1.5 g·kg-1,碱解氮126.1 mg·kg-1,全磷0.66 g·kg-1,速效磷13.4 mg·kg-1,速效钾35.2 mg·kg-1,电导率0.17 mS·cm-1,pH 7.1,土壤中砂粒(0.02~2 mm)占57.9%,粉砂粒(0.002~0.02 mm)占28.4%,黏粒( < 0.002 mm)占13.7%。
本平台为动态熏气系统,由温湿度调控系统、气体熏蒸系统、监测控制系统以及保护和报警系统共四个子系统组成。由平台附属气象站观测采集室外温度、湿度、臭氧浓度、光照、大气压力等主要气象参数信息,同时,通过温湿度传感器、光照传感器、气压测定仪和O3分析仪监测各室主要环境参数与气体浓度,通过主控系统实时接收采集信息并反馈调节各主要控制因子,使室内外环境差异维持在最小水平。本研究平台分为上下两层,下层为设备间,气体先在下层混气箱中混匀再由风机输送至上层试验区内,由于与室外空气无直接接触,臭氧浓度本底值明显低于室外环境。臭氧是以纯氧为气源,由佳环臭氧发生器(型号为QD-001-3A)产生,通过Model 49i臭氧分析仪对臭氧浓度进行即时监测(每分钟采集一次)。2015年根据试验需求,共设置2个室内对照(C-O3,室内对照)和2个高浓度臭氧处理(E-O3,设定为100 nL·L-1),熏气时间设定为每天9:00—17:00,湿度在7月22日前、后分别设定为70%、75%,温度、光照和大气压力动态模拟外界环境(图 1)。

|
| 图 1 2015年水稻生长季自然光气体熏蒸平台的控制状态 Figure 1 The performance of greenhouse-type gas fumigation chambers in 2015 rice growing season |
2015年6月15日将水稻秧苗移栽至气室,返青期内仅对温、湿度进行调控,待秧苗返青后(7月1日)开始进行熏气处理,9月4日停止熏气。平台运行期间,除因设备故障、雷雨天气以及臭氧分析仪校准等原因短时间暂停布气外(共12 d,见图 1),其余时间系统运行正常。
1.2 材料培育以目前生产上广泛选用的优质超级稻南粳9108(迟熟中粳)为供试材料。大田旱育秧,2015年5月10日播种,6月15日移栽,9月22日收获,每室12穴,两个重复,密度为27株·m-2。全生育期总施氮量为15 g·m-2,于6月10日施基肥(占总施氮量的60%),7月20日施穗肥(占总施氮量的40%);磷、钾肥总施用量均为7 g·m-2,并作为基肥一次性施用。氮肥使用尿素(含氮率46.7%)和复合肥(N-P-K:15-15-15),磷、钾肥使用复合肥。水分管理为6月15日—7月22日保持水层(约3 cm),7月23日后干湿交替,控水搁田。适时进行病虫草害防治,保证水稻正常生长发育。
1.3 测定内容和方法强、弱势粒区分:于成熟期,每个气室以6穴为一个重复分别收获,稻穗上的籽粒区分为上、下和中三个部位,即先将穗轴一分为二(一次枝梗数为奇数则上部比下部多一个一次枝梗),然后将稻穗上部一次枝梗、下部二次枝梗上的籽粒以及所有剩余籽粒分别作为稻穗上、下和中部籽粒,以此对应强势粒、弱势粒和中势粒[24-25]。
稻米氨基酸含量采用全自动氨基酸分析仪(Biochrom 30)测定。稻谷经出糙后磨粉,准确称取米粉0.100 0 g于10 mL安瓿瓶中,加入6 mol·L-1的HCl 5 mL,将安瓿瓶封口,用铝盖压实密封。用注射器抽出安瓿瓶中的空气,并用医用胶带再次封口,将安瓿瓶置于110 ℃恒温干燥箱中消化24 h后取出冷却至室温;打开安瓿瓶,过滤消化液(定量滤纸)至50 mL消化管中,室温下放置4 h;吸取2 mL滤液至10 mL试管中,用水浴锅常压蒸干去除HCl,残留物用2 mL稀释液稀释,稀释液通过0.45 μm孔径的滤膜后,即得到米粉氨基酸水解液,随后转入进样瓶中待上机测定,根据测定结果计算氨基酸含量。
1.4 统计分析方法使用Excel 2013软件进行数据处理和图表绘制,采用SPSS 19.0进行数据标准化处理和方差分析。显著水平设P < 0.01、P < 0.05、P < 0.1、P>0.1,分别用**、*、+和ns表示。
2 结果与分析 2.1 臭氧熏蒸平台的控制臭氧熏蒸期试验平台主要参数的变化示于图 1。熏蒸平台的控制一般用TAR(Target Achievement Ratio,实际测定值/设定目标值)值来表示,TAR值越接近1,则控制精度越高。结果表明,室外对照、室内对照和高浓度臭氧处理熏蒸期间平均臭氧浓度分别为61.7、18.1、100.3 nL·L-1(图 1a),臭氧精度控制的TAR值达到1.0。整个臭氧熏蒸期间的各室平均温度(图 1b)和大气压力变幅(图 1c)分别为29.3~30.8 ℃和99.8~100.3 kPa,TAR值均达到1.0。室内对照和高浓度臭氧处理整个生育期熏蒸期间相对湿度值(图 1d)分别为72.7%和72.9%,TAR均达到1.0。
2.2 臭氧浓度增加对稻穗不同部位糙米氨基酸总量、必需和非必需氨基酸总量的影响臭氧浓度增加对南粳9108稻穗各部位糙米氨基酸总量的影响如图 2所示。该品种全穗糙米氨基酸总量平均(两臭氧处理平均)为62.7 mg·g-1,稻穗上部、中部和下部氨基酸总量分别为61.7、64.2、62.4 mg·g-1,中部最大,但无显著差异。与对照相比,臭氧处理使该品种糙米氨基酸总量平均增加5.5%,达显著水平。从稻穗不同部位看,臭氧浓度增加使稻穗上部、中部和下部糙米氨基酸总量分别增加1.6%、2.9%和12.3%,稻穗中、下部均达极显著或显著水平。方差分析表明,臭氧处理与稻穗不同部位对糙米氨基酸总量有微弱的互作效应。

|
| 图 2 臭氧浓度增加对南粳9108稻穗不同部位糙米氨基酸总量、必需和非必需氨基酸总量的影响 Figure 2 Effect of ozone stress on total of essential, nonessential and all amino acids in brown rice at different positionsonapanicle of Nanjing 9108 **:P < 0.01, *: P < 0.05, ns: 不显著, no significance。下同 |
水稻糙米氨基酸总量为必需氨基酸总量和非必需氨基酸总量之和。图 2表明,糙米必需、非必需氨基酸的总量平均(两臭氧处理平均)分别为21.5、41.3 mg·g-1,后者为前者的两倍,但两参数不同部位间均无显著差异。与对照相比,臭氧处理使南粳9108必需氨基酸、非必需氨基酸总量平均(两臭氧处理平均)分别增加5.4%和5.6%,后者达0.01显著水平。从部位看,臭氧浓度增加使稻穗上部、中部和下部糙米必需氨基酸总量分别增加1.6%、3.1%和11.8%,非必需氨基酸总量分别增加1.7%、2.8%和12.5%,稻穗上部未达显著水平,但稻穗中部和下部均达显著水平。方差分析表明,臭氧处理与稻穗不同部位对糙米非必需氨基酸总量有显著的互作效应。
2.3 臭氧浓度增加对稻穗不同部位糙米必需、非必需氨基酸占氨基酸总量百分比的影响图 3为臭氧浓度增加对稻米必需、非必需氨基酸占氨基酸总量百分比的影响。稻米必需氨基酸占氨基酸总量的百分比(平均34.2%)明显小于非必需氨基酸占氨基酸总量的百分比(平均65.8%),但两指标稻穗不同部位间均无差异。臭氧处理对稻穗不同部位糙米必需或非必需氨基酸占氨基酸总量的百分比均没有影响。方差分析表明,臭氧处理与稻穗部位间的互作对两参数均无显著影响。

|
| 图 3 臭氧浓度增加对南粳9108稻穗不同部位糙米必需和非必需氨基酸占氨基酸总量百分比的影响 Figure 3 Effect of ozone stress on the percentage of essential or nonessential amino acids to total amino acids in brown rice at different positionsona panicle of Nanjing 9108 |
组成蛋白质的一部分氨基酸必须由食物供给(而非人体合成),称为必需氨基酸。臭氧浓度增加对水稻糙米中7种必需氨基酸含量的影响列于表 1。稻穗上部、中部和下部糙米蛋氨酸含量平均分别为1.02、1.09、0.89 mg·g-1,部位间差异达0.1显著水平;除蛋氨酸外,糙米中其他6种必需氨基酸含量的部位差异均未达显著水平。与对照相比,臭氧浓度增加使水稻糙米中所有必需氨基酸含量均呈增加趋势,其中苏氨酸(+10.2%)、亮氨酸(+7.8%)均达显著水平。从着生部位看,臭氧浓度增加条件下稻穗各部位诸必需氨基酸含量多呈增加趋势,其中稻穗上部氨基酸含量最高增幅为6.7%,但均未达显著水平;稻穗中部最高增幅为9.9%,有4个必需氨基酸达0.1显著水平;稻穗下部所有必需氨基酸含量均增加,最高增幅达19.2%,除蛋氨酸外,均达0.1以上显著水平。方差分析表明,臭氧处理与部位对糙米苯丙氨酸有显著的互作效应。

|
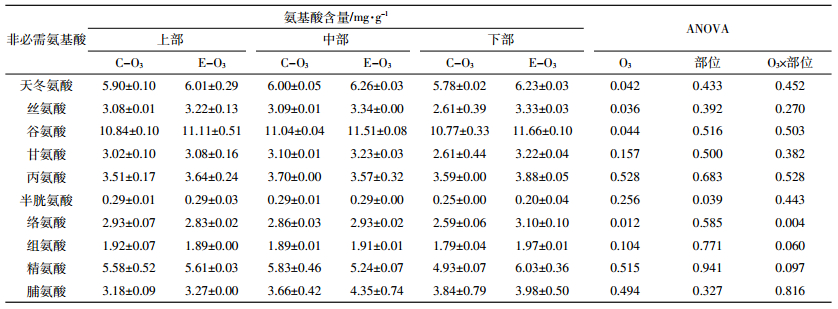
组成蛋白质的另一部分氨基酸可能由人体自身合成,称为非必需氨基酸。表 2结果表明,稻穗上部、中部和下部糙米半胱氨酸含量平均分别为0.29、0.29、0.23 mg·g-1,部位间差异达显著水平;除半胱氨酸外,糙米中其他9种非必需氨基酸含量的部位差异均未达显著水平。与对照相比,臭氧浓度增加使水稻糙米中所有非必需氨基酸(半胱氨酸除外)含量均呈增加趋势,其中天冬氨酸(+4.7%)、丝氨酸(+12.6%)、谷氨酸(+5.0%)和络氨酸(+5.7%)增幅达显著水平,甘氨酸(+9.0%)、组氨酸(+3.0%)接近0.1显著水平。从着生部位看,臭氧浓度增加条件下稻穗各部位诸非必需氨基酸含量多呈增加趋势,其中稻穗上部、中部和下部最高增幅分别为4.5%、19.0%、27.7%,稻穗上部所有非必需氨基酸均未达显著水平,但稻穗中部、下部多数达到或接近0.1以上显著水平。方差分析表明,臭氧处理与部位对糙米络氨酸、组氨酸和精氨酸的影响分别达0.01、0.1和0.1显著水平。
|
本试验依托的自然光气体熏蒸平台在借鉴传统气室优点的基础上增大了试验空间,采用自然采光特别是土培方式培育水稻植株,并动态追踪室外的空气温度。水稻生长季平台运行数据表明,自然光气体熏蒸平台控制准确,满足试验要求(图 1)。
作为重要的稻米营养品质指标,臭氧浓度增加对稻米氨基酸含量的报道甚少。仅有的一例水稻研究表明,臭氧浓度增加24.5%使杂交籼稻汕优63稻米氨基酸、必需和非必需氨基酸总量平均增加12%~14%[17]。本研究采用的超级稻南粳9108为常规粳稻,结果表明,100 nL·L-1臭氧浓度使其稻米氨基酸、必需和非必需氨基酸总量显著增加(图 2),但平均增幅(+5%~6%)小于敏感的杂交籼稻汕优63[17]。这一结果证实,臭氧浓度增加对稻米氨基酸的影响亦可能存在基因型差异。稻米蛋白质的质量取决于氨基酸的平衡。由于臭氧浓度增加条件下稻米必需和非必需氨基酸总量的增幅相近,对稻米必需或非必需氨基酸占氨基酸总量的百分比无显著影响(图 2、图 3),这一结果与汕优63的表现相同[17],说明高浓度臭氧环境下稻米氨基酸的浓度呈增加趋势,但氨基酸的平衡无明显变化。
关于臭氧浓度增加对稻米各种氨基酸组分的影响,Zhou等[17]FACE研究表明,臭氧胁迫使杂交籼稻汕优63稻米15种氨基酸显著增加。本研究在此基础上多测定了半胱氨酸和络氨酸这两种氨基酸,结果表明,除半胱氨酸外,臭氧胁迫使糙米中16种氨基酸的含量均呈增加趋势,其中3个必需氨基酸(苏氨酸、亮氨酸和苯丙氨酸)、4个非必需氨基酸(丝氨酸、谷氨酸、络氨酸和天冬氨酸)的增幅均达显著水平,平均最高增幅为12.6%(表 1~表 2)。这一结果小于已报道的杂交籼稻汕优63[17],但大于Fuhrer等[26]对小麦的研究结果。Fuhrer等[26]研究发现,尽管高臭氧浓度(比环境空气高2.5倍)使谷粒蛋白质浓度增加,但谷粒中的16种氨基酸浓度在各种臭氧熏蒸浓度下均无显著变化。
一般认为,臭氧熏蒸导致植物食用部分蛋白质或氨基酸浓度增加的原因可能有两个:第一,与“浓缩效应”有关,即碳代谢途径受臭氧胁迫的影响可能大于氮代谢;第二,与臭氧胁迫导致植株的早衰有关,后者有利于灌浆期蛋白质向谷粒的转移[7]。本试验同期测定结果表明,臭氧胁迫使南粳9108粒重显著下降,但抽穗和成熟的日期均没有变化,说明臭氧熏蒸使稻米氨基酸浓度增加可能只与“浓缩效应”有关,当然这还需要更多的研究验证。尽管臭氧浓度增加使稻米氨基酸浓度增加,但由于臭氧导致产量大幅下降(平均下降28.9%),所以单位面积的氨基酸产量大幅下降。
前期研究发现,臭氧浓度增加对稻米蛋白质性质的影响因臭氧熏蒸强度[10]、环境O3水平[13]以及供试品种[10, 27]而异,但同一品种稻穗内部强、弱势粒间是否存在差异未见报道。为此,本试验根据籽粒在稻穗上的着生位置区分为上、中和下三个部位,分别对应强势粒、中势粒和弱势粒[24-25]。稻米氨基酸及其组分浓度的测定数据表明,多数情形下,臭氧浓度增加对这些参数影响表现为下部>中部>上部(图 2、表 1、表 2)。例如,臭氧浓度增加对稻穗上部测定的所有17种氨基酸含量均无显著影响,而稻穗中部、下部分别有8种、11种氨基酸浓度的响应达到0.1以上的显著水平,稻穗上、中和下部稻米诸氨基酸浓度的最高增幅分别为7%、19%和28%。方差分析结果显示,臭氧与部位对部分测定指标有显著的互作效应(表 1、表 2)。笔者认为,上述这种趋势可能与强、弱势粒的灌浆时间不同有关:与稻穗上部的强势粒相比,位于稻穗下部的弱势粒灌浆明显慢、灌浆时间长,因此其籽粒生长过程受臭氧浓度增加的伤害更大,导致其体积和重量的降幅均显著大于强势粒,“浓缩效应”表现得更为明显。水稻强、弱势粒对环境响应的这种差异说明,开展臭氧浓度增加与稻米品质的研究需要区分籽粒在稻穗上的着生部位甚至粒位。本试验研究还发现,稻米不同氨基酸对臭氧浓度增加的响应存在差异,这可能与臭氧浓度增加对诸氨基酸合成过程的影响程度不同有关,例如,合成过程的有关酶类尚待进一步探明。结合其他研究,弱势粒结实和品质指标对环境变化的响应通常大于强势粒[18],从另一侧面说明了它的可调性。这些现象为通过作物管理技术的改进并进一步挖掘水稻生产潜力提供了依据。
前期研究表明臭氧浓度增加对水稻生长、产量及品质的影响多受供试品种的影响[7, 9],本试验只观察了一个超级稻品种稻米相关氨基酸性状的变化,下一步还需开展不同类型水稻,特别是不同穗型水稻的大田实验研究,并从植株碳氮代谢响应角度解析其生理机制。
4 结论本文以生产上大面积使用的超级稻品种南粳9108为试验对象,首次研究了臭氧浓度增加对稻米氨基酸含量的影响及其与稻穗着生部位的关系。臭氧浓度增加使供试品种糙米氨基酸总量、必需和非必需氨基酸总量、除半胱氨酸外的16种必需和非必需氨基酸含量均呈增加趋势,但稻穗中、下部籽粒的响应明显大于稻穗上部籽粒。与此不同,臭氧浓度增加对该品种氨基酸的平衡无显著影响。以上结果说明臭氧浓度增加使超级稻南粳9108糙米中各种氨基酸组分的含量均呈上升趋势,且多数情况下,弱势粒的增幅大于强势粒。
| [1] | Emberson L D, Ashmore M R, Murray F, et al. Impacts of air pollutants on vegetation in developing countries[J]. Water, Air, and Soil Pollution, 2001, 130(1/2/3/4): 107–118. |
| [2] | Fowler D, Amann M, Anderson F, et al. Ground-level ozone in the 21st century:Future trends, impacts and policy implications[M]. London: The Royal Society, 2008. |
| [3] | Krupoa S, McGrath M T, Andersen C P, et al. Ambient ozone and plant health[J]. Plant Disease, 2001, 85(1): 4–12. DOI:10.1094/PDIS.2001.85.1.4 |
| [4] | Feng Z Z, Kobayashi K. Assessing the impacts of current and future concentrations of surface ozone on crop yield with meta-analysis[J]. Atmospheric Environment, 2009, 43(8): 1510–1519. DOI:10.1016/j.atmosenv.2008.11.033 |
| [5] | Ainsworth E A. Rice production in a changing climate:A meta-analysis of responses to elevated carbon dioxide and elevated ozone concentration[J]. Global Change Biology, 2008, 14(7): 1642–1650. DOI:10.1111/j.1365-2486.2008.01594.x |
| [6] | Ainsworth E A, McGrath J M. Direct effects of rising atmospheric carbon dioxide and ozone on crop yields[M]//Lobell D, Burke M. Climate change and food security:Advances in Global Change Research. 37. Berlin:Springer Netherlands, 2010:109-130. |
| [7] | Wang Y X, Michael F. Stressed food-The impact of abiotic environmental stresses on crop quality[J]. Agriculture, Ecosystems and Environment, 2011, 141: 271–286. DOI:10.1016/j.agee.2011.03.017 |
| [8] | Sun J D, Yang L X, Wang Y L, et al. FACE-ing the global change:Opportunities for improvement in photosynthetic radiation use efficiency and crop yield[J]. Plant Science, 2009, 177(6): 511–522. DOI:10.1016/j.plantsci.2009.08.003 |
| [9] | 杨连新, 王余龙, 石广跃, 等. 近地层高臭氧浓度对水稻生长发育影响研究进展[J]. 应用生态学报, 2008, 19(4): 901–910. YANG Lian-xin, WANG Yu-long, SHI Guang-yue, et al. Responses of rice growth and development to elevated near-surface layer ozone (O3) concentration:A review[J]. Chinese Journal of Applied Ecology, 2008, 19(4): 901–910. |
| [10] | Frei M, Kohno Y, Tietze S, et al. The response of rice grain quality to ozone exposure during growth depends on ozone level and genotype[J]. Environmental Pollution, 2012, 163(4): 199–206. |
| [11] | Wang Y X, Yang L X, Han Y, et al. The impact of elevated tropospheric ozone on grain quality of hybrid rice:A free-air gas concentration enrichment (FACE) experiment[J]. Field Crops Research, 2012, 129(1): 81–89. |
| [12] | 宋琪玲, 齐义涛, 赵轶鹏, 等. 自由空气中臭氧浓度升高对"武运粳21"稻米物性及食味品质的影响[J]. 中国生态农业学报, 2013, 21(5): 566–571. SONG Qi-ling, QI Yi-tao, ZHAO Yi-peng, et al. Impact of free air ozone concentration enrichment on cooked rice (Wuyunjing21) texture and palatability[J]. Chinese Journal of Eco-Agriculture, 2013, 21(5): 566–571. DOI:10.3724/SP.J.1011.2013.00566 |
| [13] | Wang Y X, Song Q L, Frei M, et al. Effects of elevated ozone, carbon dioxide, and the combination of both on the grain quality of Chinese hybrid rice[J]. Environmental Pollution, 2014, 189(12): 9–17. |
| [14] | Sarkar A, Agrawal S B. Evaluating the response of two high yielding Indian rice cultivars against ambient and elevated levels of ozone by using open top chambers[J]. Journal of Environmental Management, 2012, 95(2): S19–S24. |
| [15] | 郭建平, 王春乙, 温民, 等. 大气中O3浓度变化对水稻影响的试验研究[J]. 作物学报, 2001, 27(6): 822–826. GUO Jian-ping, WANG Chun-yi, WEN Min, et al. The experimental study on the impact of atmospheric O3 variation on rice[J]. Acta Agronomica Sinica, 2001, 27(6): 822–826. |
| [16] | Huang Y Z, Sui L H, Wang W, et al. Visible injury and nitrogen metabolism of rice leaves under ozone stress, and effect on sugar and protein contents in grain[J]. Atmospheric Environment, 2012, 62: 433–440. DOI:10.1016/j.atmosenv.2012.09.002 |
| [17] | Zhou X D, Zhou J, Wang Y X, et al. Elevated tropospheric ozone increased grain protein and amino acid content of a hybrid rice without manipulation by planting density[J]. Journal of the Science of Food and Agriculture, 2015, 95(1): 72–78. DOI:10.1002/jsfa.2015.95.issue-1 |
| [18] | 杨建昌. 水稻弱势粒灌浆机理与调控途径[J]. 作物学报, 2010, 36(12): 2011–2019. YANG Jian-chang. Mechanism and regulation in the filling of inferior spikelets of rice[J]. Acta Agronomica Sinica, 2010, 36(12): 2011–2019. |
| [19] | 袁隆平. 选育超高产杂交水稻的进一步设想[J]. 杂交水稻, 2012, 27(6): 1–2. YUAN Long-ping. Conceiving of breeding further super-high-yield hybrid rice[J]. Hybrid Rice, 2012, 27(6): 1–2. |
| [20] | 李建武, 张玉烛, 吴俊, 等. 超高产水稻新组合Y两优900百亩方15.40 t/hm2高产栽培技术研究[J]. 中国稻米, 2014, 20(6): 1–4. LI Jian-wu, ZHANG Yu-zhu, WU Jun, et al. High-yielding cultural techniques of super hybrid rice YLY 900 yielded 15.40 t/hm2 on a 6. 84 hm2 scale[J]. China Rice, 2014, 20(6): 1–4. |
| [21] | Shi G Y, Yang L X, Wang Y X, et al. Impact of elevated ozone concentration on yield of four Chinese rice cultivars under fully open-air field conditions[J]. Agriculture, Ecosystems and Environment, 2009, 131(3/4): 178–184. |
| [22] | 彭斌, 赖上坤, 李潘林, 等. 开放式空气中臭氧浓度升高对超级稻Ⅱ优084生长和产量的影响[J]. 农业环境科学学报, 2014, 33(2): 217–223. PENG Bin, LAI Shang-kun, LI Pan-lin, et al. Effects of free air ozone enrichment on growth and yield of super riceⅡ-You 084[J]. Journal of Agro-Environment Science, 2014, 33(2): 217–223. |
| [23] | 赵轶鹏, 邵在胜, 宋琪玲, 等. 一种新型自然光气体熏蒸平台:系统结构与控制精度[J]. 农业环境科学学报, 2012, 31(11): 2082–2093. ZHAO Yi-peng, SHAO Zai-sheng, SONG Qi-ling, et al. System structure and control accuracy of a solar-illuminated gas fumigation platform[J]. Journal of Agro-Environment Science, 2012, 31(11): 2082–2093. |
| [24] | 张庆, 王娟, 景立权, 等. 叶面施用不同形态锌化合物对稻米锌浓度及有效性的影响[J]. 中国水稻科学, 2015, 29(6): 610–618. ZHANG Qing, WANG Juan, JING Li-quan, et al. Effect of foliar application of different Zn compounds on Zn concentration and bioavailability in brown rice[J]. Chinese Journal of Rice Science, 2015, 29(6): 610–618. |
| [25] | 周三妮.不同条件下结实期叶面施锌对稻米锌浓度及有效性的影响[D].扬州:扬州大学, 2015:1-91. ZHOU San-ni. The effects of foliar Zn application during grain filling stage on Zn concentration and bioavailability under different growth conditions[D]. Yangzhou:Yangzhou University, 2015:1-91. |
| [26] | Fuhrer J, Lehnherr B, Moeri P B, et al. Effects of ozone on the grain composition of spring wheat grown in open-top chambers[J]. Environmental Pollution, 1990, 65(2): 181–192. DOI:10.1016/0269-7491(90)90183-D |
| [27] | Jing L Q, Dombinov V, Shen S B, et al. Physiological and genotype-specific factors associated with grain quality changes in rice exposed to high ozone[J]. Environmental Pollution, 2016, 210: 397–408. DOI:10.1016/j.envpol.2016.01.023 |