 2019, Vol. 38
2019, Vol. 38



2. 海峡两岸红壤区水土保持协同创新中心, 福州 350002;
3. 南方红壤区水土保持国家林业局重点实验室, 福州 350002
2. Co-innovation Center for Soil and Water Conservation in Red Soil Region of the Cross-straits, Fuzhou 350002, China;
3. Key Laboratory of the State Forestry Administration for Soil and Water Conservation in Southern Red Soil Region, Fuzhou 350002, China
矿山开采废弃地的污染及其治理是我国及许多发展中国家目前面临的主要环境问题之一,不同类型矿山废弃地植被恢复的限制因子存在一定差异,其中土壤酸度低和重金属污染是大多金属矿山废弃地植被恢复的主要限制因子[1-3]。福建长汀的稀土资源储量为福建省之最,达13.2万t,由于稀土开采长期处于工艺落后、管理不善等状况,使矿区土壤结构遭到严重破坏,不仅造成水土流失,还造成重金属污染与土壤酸化[4]。研究表明长汀稀土矿区废弃堆浸池的土壤酸度为4.05,重金属Cd含量为7.64 mg·kg-1,Cd综合生态风险指数达极重污染水平,酸和Cd是限制稀土矿废弃地植被恢复的重要限制因子[5]。
宽叶雀稗(Paspalum wettsteinii)是禾本科多年生草本植物,其茎下部贴地面呈匍匐状,分蘖能力强,着地部分节上可长出不定根,生长速度快,在干旱贫瘠的红、黄壤坡地亦能生长,具有抗旱性强、耐酸、耐贫瘠等特点[6],在长汀稀土矿废弃地治理中有很好的植被恢复效果,其与胡枝子、木荷、枫香和山杜英搭配的植被恢复模式可使土壤容重降低到1.33~1.35 g · cm-3,达到土壤良性结构的要求,同时土壤肥力指数可达到0.544,对土壤肥力有较好的改良效果,得到了广泛应用[7]。在长汀稀土矿废弃地治理过程中采取撒播宽叶雀稗种子的方法进行植被恢复,说明其种子对酸和Cd胁迫具有较强的耐性,其中可能存在一些有待挖掘的特殊响应机理。
鉴于此,本研究以宽叶雀稗种子为试验材料,设置不同酸和Cd胁迫处理,在人工气候箱中开展宽叶雀稗种子对酸和Cd胁迫的响应研究,测定不同胁迫处理条件下宽叶雀稗种子萌发、幼苗生长、生物量等指标,以及质膜完整性和亚细胞分布,比较不同胁迫处理下宽叶雀稗种子萌发之间的差异,分析宽叶雀稗种子在不同酸和Cd胁迫下的响应,以期为稀土矿废弃地及同类矿山废弃地的生态恢复提供科技支撑。
1 材料与方法 1.1 试验材料宽叶雀稗种子购买于深圳市鑫淼森生态环境建设有限公司,种子在4 ℃冰箱中保存。酸胁迫液采用CH3COOH进行配制,Cd胁迫液采用分析纯试剂Cd(CH3CO2)2进行配制。
1.2 试验方法宽叶雀稗种子先用0.3%的高锰酸钾溶液浸泡30 min,然后用纯水冲洗数遍后室温浸泡24 h。酸胁迫的pH值分别为3.5、4.5、5.5,Cd胁迫浓度分别为5、10、20 mg·L-1(Cd5、Cd10、Cd20),同时设置纯水处理作为对照(CK,pH值为6.1),每个处理均3个重复。利用培养皿进行种子萌发胁迫实验,每个培养皿(9 cm)放两张滤纸,并各加5 mL的胁迫液,选取大小一致、色泽鲜艳、饱满的宽叶雀稗种子,每个处理整齐排列50粒。试验期间采用称重法保持培养皿湿度,并每2 d更换一次滤纸和胁迫液,以排除水分蒸发和胁迫液浓度变化对实验的影响。试验在人工气候箱中进行,培养条件为温度25 ℃,湿度75%,光照强度10 000 lx,昼/夜光照时间14 h/10 h。以种子胚根突破种皮,肉眼可见为萌发标志,每天记录发芽数,以连续5 d萌发数保持一致为发芽结束标志。试验第4 d统计发芽势,第18 d统计发芽率。
1.3 测定指标 1.3.1 萌发指标测定[8]发芽率=萌发种子数/种子总数×100%
发芽势=发芽高峰期发芽的种子数/种子总数× 100%
发芽指数(GI)=Σ(Gt/Dt)
活动指数(VI)=GI×g
式中:Gt为t时间内的萌发数;Dt为相应的萌发时间,d;g为芽长度+根长度。
1.3.2 萌发抑制指数[8]抑制指数(RI)=(对照指标值-处理指标值)/对照指标值
当RI=0时,酸、Cd胁迫对种子萌发无明显作用;当RI>0时,酸、Cd胁迫对种子萌发起抑制作用;当RI < 0时,酸、Cd胁迫对种子萌发起促进作用。
1.3.3 胚根长、胚芽长的测定萌发结束后,参考鲁如坤[9]的方法测量根长、芽长。在每盘培养皿中各选取20株长势一致的幼苗,用游标卡尺测量其根长、芽长。
1.3.4 生物量的测定萌发结束后,用四位电子天平分别称量根、芽和胚的鲜质量;然后105 ℃杀青30 min,80 ℃烘干至恒质量,用四位电子天平称量其干质量。
1.3.5 测定质膜完整性质膜完整性检验采用Evans blue染色的方法[10]。从酸或Cd胁迫后的培养皿里分别取长度为2 cm左右的宽叶雀稗幼苗根尖,每个浓度3~5根,放于4 ℃冷处理的0.5 mmol·L-1的CaCl2(pH 4.5)溶液中浸泡5 min,然后用蒸馏水洗净吸干,再将根浸泡于4 mL伊文思蓝溶液(0.025%伊文思蓝溶于100 μmol·L-1 pH 5.6的CaCl2溶液中)中30 min,取出后用蒸馏水冲洗,最后在蒸馏水中浸泡15 min,然后在光学显微镜下观察拍照,以测定原生质膜的完整性。
1.3.6 叶片超显微结构的测定将样品放入2.5%的戊二醛溶液(用pH 7.4的磷酸缓冲溶液配制)中,并于4 ℃条件下固定过夜。次日用0.1 mol·L-1磷酸缓冲溶液(pH 7.4)清洗4次,将洗涤过的样品放入1%锇酸溶液(磷酸缓冲溶液配制)中4 ℃浸泡过夜,再次用磷酸缓冲溶液(pH 7.4)清洗4次。然后用乙醇逐级脱水(50%、70%、80%、90%、100%),再用丙酮置换2次。Spur包埋,将样品放入盛有纯包埋剂的包埋板中,再将包埋板置于65 ℃条件下各聚合48 h以上。用EM UC7型超薄切片机进行切片,切片厚度为70 nm,然后分别用醋酸双氧铀、醋酸铅各染色10 min,清洗后使用Tecnai G2 Spirit型透射电子显微镜对宽叶雀稗胚芽组织进行检测。并采用能谱仪(EDS)对样品重金属含量进行进一步定性分析。
1.4 数据处理采用Excel 2013和SPSS 19.0对数据进行统计分析,使用单因素方差分析(One-way ANOVA)进行差异显著性分析,用Duncan′s法进行多重比较,显著性水平为0.05,图表中数据均用3个重复的平均值±标准差表示,采用Origin 8.5作图。
2 结果与分析 2.1 酸、Cd胁迫对宽叶雀稗种子萌发指标的影响由表 1可知,随着pH值的降低,发芽指标均表现为先增后减趋势,pH 5.5处理和pH 4.5处理的发芽势、发芽指数和活力指数与对照均无显著差异(P> 0.05),pH 3.5处理则显著小于对照(P < 0.05)。pH 4.5处理的发芽率显著大于对照(P < 0.05),为85.33%,是对照的1.11倍;pH 3.5处理的发芽率与对照无显著差异(P>0.05)。随着Cd浓度的升高,发芽指标均呈现先增后减再增的趋势,各胁迫浓度下的发芽率、发芽势和发芽指数与对照均无显著差异(P>0.05),但其活力指数均显著小于对照(P < 0.05),且表现为CK>Cd 5> Cd 20>Cd 10。
|
|
表 1 酸、Cd胁迫下宽叶雀稗种子萌发指标的变化 Table 1 Changes in germination indexs of Paspalum wettsteinii seeds under acid and Cd stresses |
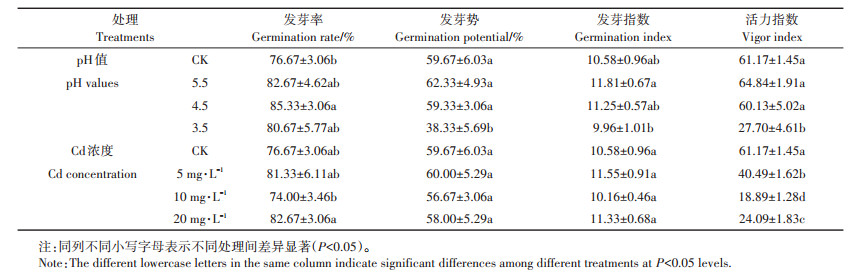
酸、Cd胁迫下宽叶雀稗根长、芽长和长度根冠比的变化见图 1。在酸胁迫下,pH 5.5和pH 4.5处理下的根长、芽长和长度根冠比均与对照无显著差异(P> 0.05),且根长均大于芽长;pH 3.5处理下各指标均显著小于其他处理组(P < 0.05),根长、芽长分别是对照的28%和72%,且根冠比小于1,说明pH 3.5严重影响根长、芽长的生长,且抑制作用表现为根>芽。在Cd胁迫下,随着Cd浓度的增大,根长、芽长均呈减小趋势,CK组的根长、芽长显著大于其他处理组(P < 0.05),表现为CK>Cd 5>Cd 20>Cd 10,且各处理的根冠比均大于1。

|
不同小写字母表示同一指标在不同处理间差异显著(P<0.05) The different lowercase letters indicate that the same index varies significantly among different treatments at P<0.05 levels 图 1 酸、Cd胁迫下宽叶雀稗幼苗根长、芽长和长度根冠比 Figure 1 Root length, bud length and root-shoot ratio of the seedlings under acid and Cd stresses |
由表 2可知,pH 5.5处理对各萌发指标均有促进效果,同时促进芽生长且不抑制根长;pH 4.5对发芽率、芽长的促进效果为酸处理中最大,但会抑制发芽势、活力指数和根长;pH 3.5处理下,除发芽率外的其他指标均受到抑制,尤其是根长、活力指数。由此可知,随着pH值的减小,抑制作用逐渐增大。
|
|
表 2 酸、Cd胁迫下宽叶雀稗萌发指标、根长和芽长的抑制指数 Table 2 Inhibitory index of germination index, root length and bud length under acid and Cd stress |
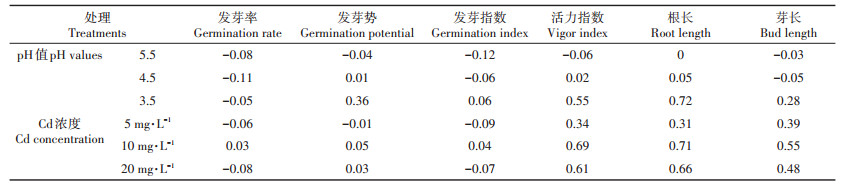
宽叶雀稗在Cd胁迫下,其活力指数、根长和芽长均受到抑制,其抑制作用表现为Cd 10>Cd 20 >Cd 5,且在Cd 10和Cd 20处理下,抑制作用表现为根长>芽长。Cd 5处理会促进种子的发芽率、发芽势和发芽指数,而Cd 10处理则相反;Cd 20处理对发芽率的促进作用为各处理中最大。说明相较于其他Cd胁迫浓度,5 mg·L-1较适合宽叶雀稗种子萌发和幼苗生长,10 mg·L-1对其的抑制性最大。
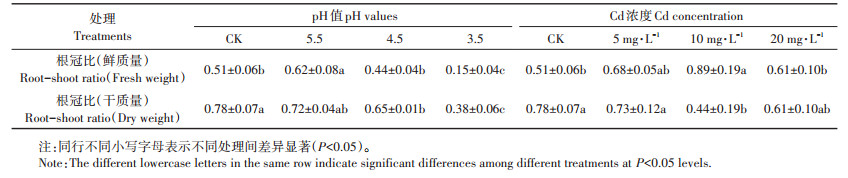
2.4 酸、Cd胁迫对宽叶雀稗生物量的影响植物的生长环境会直接或间接地影响植物的生长发育,植物的鲜质量和干质量是衡量植物生长情况的一个重要指标,其鲜质量根冠比和干质量根冠比的大小反映了植物地下部分与地上部分的相关性[11]。由图 2A、图 2B和表 3可知,随着pH值的减小,根鲜质量、干质量和芽干质量均呈减小趋势,芽鲜质量和胚鲜质量、干质量则相反;pH 3.5处理下的根干质量、芽干质量均显著小于其他处理组(P < 0.05),pH 5.5和pH 4.5处理下的根干质量和芽干质量均与对照无显著差异(P>0.05),但是pH 4.5的芽干质量显著大于pH 5.5(P < 0.05),是其1.07倍;pH 3.5的鲜质量、干质量根冠比显著小于对照(P < 0.05)。随着Cd浓度的升高,根和芽的鲜质量、干质量均呈减小趋势(图 2C、图 2D和表 3),CK的根和芽的鲜质量、干质量均显著大于其他各处理组(P < 0.05),其根干质量、芽干质量分别是Cd 5、Cd 10和Cd 20的2.54、7.06、6.31倍和2.61、5.18、5.32倍;Cd 5处理下的胚鲜质量、干质量均显著小于对照(P < 0.05);Cd 10的干质量根冠比也显著小于对照(P < 0.05)。

|
不同小写字母表示植物某部分的鲜质量或干质量在不同处理间差异显著(P<0.05) The different lowercase letters indicate that the fresh weight or dry weight of a part of the plant is significantly different among different treatments at P<0.05 levels 图 2 酸、Cd胁迫下宽叶雀稗幼苗各部分鲜质量、干质量的变化 Figure 2 Changes of fresh weight and dry weight in different parts of the seedling under acid and Cd stresses |
|
|
表 3 酸、Cd胁迫对宽叶雀稗幼苗根冠比(鲜质量和干质量)的影响 Table 3 Effects of acid and Cd stresses on root-shoot ratio(fresh weight and dry weight)of the seedlings |
通过用伊文思蓝染色法对宽叶雀稗根尖进行染色分析发现(图 3),对于酸胁迫而言,染色的着色程度由浅到深为pH 5.5 < CK(pH 6.1) < pH 4.5 < pH 3.5,说明pH 3.5处理严重损害了根尖质膜;对于Cd胁迫而言,染色的着色程度由浅到深为CK < Cd 5 < Cd 20 < Cd 10,说明Cd胁迫均会对根尖质膜造成一定程度的损害,其中Cd 10处理最为严重。

|
图 3 酸、Cd胁迫下宽叶雀稗幼苗根尖质膜完整性 Figure 3 The integrity of plasmalemma in the root tip cells under acid and Cd stresses |
由芽亚细胞结构图(图 4)可知,在CK组(图 4A、图 4B)中,芽细胞无质壁分离现象,核糖体形态完整,叶绿体紧贴细胞边缘,部分叶绿体外膜变薄,有的断裂,形成髓样结构,但其内基粒和类囊体结构清晰,排列整齐,淀粉粒数量较多。在酸胁迫下,pH 5.5处理(图 4C、图 4D)的宽叶雀稗芽细胞形态完整,细胞壁和细胞膜膜质平滑,细胞内各细胞器结构完好,叶绿体周围有少量髓样结构,与CK形态相似;pH 4.5处理(图 4E、图 4F)的细胞间距变大,出现轻微质壁分离,部分细胞的叶绿体变形,趋于圆形,且不再贴壁分布,外被膜断裂、模糊,出现较多髓样结构,基粒、类囊体结构模糊;pH 3.5处理(图 4G、图 4H)的细胞质基质不均匀,叶绿体内膜模糊,有的破裂,基质外流,部分淀粉粒肿胀、断裂,且质体小球变大、增多,线粒体发生变形,外被膜部分断裂不完整,内部基质外渗,管状嵴分布不均匀,数量减少甚至消失不见,细胞受到较大损伤。

|
A,B-CK;C,D-pH 5.5;E,F-pH 4.5;G,H-pH 3.5;CW:细胞壁Cell wall;CM:细胞膜Cell membrane;Ch:叶绿体Chloroplast;P:质体小球Plastoglobuli;SG:淀粉粒Starch grains;Thy:类囊体Thylakoid;R:核糖体Ribosome;MS:髓样结构Myelinic structure;Mi:线粒体Mitochondrion。下同The same below 图 4 酸胁迫下宽叶雀稗幼苗芽亚细胞结构 Figure 4 Effects of acid stress on bud subcellular structure of the seedlings |
Cd胁迫下的芽亚细胞结构变化情况见图 5。Cd 5处理(图 5I、图 5J)细胞质开始脱离细胞壁,部分细胞的细胞结构较完整,叶绿体淀粉粒数量增多,质体小球变大,但部分细胞出现叶绿体等细胞器解体现象,且叶绿体、线粒体和细胞壁周围出现少量黑色颗粒;Cd 10胁迫(图 5K、图 5L)对芽细胞的损伤严重,表现为细胞壁出现断裂,细胞膜变薄,细胞基质大量外流,叶绿体、细胞核等细胞器出现解体,内质网和高尔基体等细胞器空泡化或消失,细胞出现空泡化现象,且细胞壁周围聚积较多黑色颗粒,细胞受损非常严重;Cd 20胁迫(图 5M、图 5N)芽细胞间距变大,细胞膜变薄,细胞器受损,细胞质基质不均匀,叶绿体外膜破裂,基质外流,基粒排列混乱,类囊体变形,细胞壁和叶绿体间积聚很多黑色颗粒。

|
A, B-CK;I,J-Cd 5;K,L-Cd 10;M,N-Cd 20 图 5 Cd胁迫下宽叶雀稗幼苗芽亚细胞结构 Figure 5 Effects of Cd stress on bud subcellular structure of the seedlings |
对以上Cd胁迫下的芽亚细胞(图 5中圆圈部位)进行能谱分析(图 6)发现,Cd 5处理和Cd 20处理的芽细胞能谱图中,Cd、Cl、Al含量均较多;Cd 10处理的芽细胞能谱图中Cd、Cl含量较多,且在不同浓度下,Cd均主要吸附于3~4 keV位点间,可证实存在于细胞壁、叶绿体周围的黑色颗粒物的主要成分是重金属Cd。

|
I,J-Cd 5;K,L-Cd 10;M,N-Cd 20 图 6 不同Cd浓度胁迫下宽叶雀稗幼苗芽细胞能谱图 Figure 6 Energy spectra of the seedling bud cells under different Cd concentration stress |
不同的植物生长习性不同,其适宜生长的土壤酸碱度也会有所差异。土壤酸化或碱化都会影响土壤的物理、化学甚至生物学特性,继而会对植物吸收土壤中水分和养分产生影响,最终影响植物正常的生长发育[11]。有研究表明[12-14],油菜根系的生长发育受土壤酸碱变化的影响,在pH 6.1时油菜的根系活力最大,随着pH值的减小,根系长度、根体积和根系生物量均会减小;香根草可在pH 4.0~9.0的土壤环境中生长良好,对土壤酸碱度的适应范围较广;柳枝稷在pH >7.0的土壤环境中生长较好,应尽量避免在土壤pH < 5.0的地区种植。种子保持活力并萌发成苗对植物种群的繁衍和生存起着决定作用,同时也决定着植物进入自然和生态系统的时间[15]。种子发芽势、发芽指数均是表示种子活力的重要指标,发芽势的高低表示种子活力的强弱,发芽势高的种子发芽整齐且出苗一致,种子的活力程度受种子萌发环境的影响[16]。本研究发现酸胁迫会促进种子萌发,pH 4.5~6.1处理对宽叶雀稗的生物量影响较小;pH 4.5处理下其发芽率最高,但对其他指标会产生抑制作用;pH 5.5处理对各萌发指标均有促进作用,且促进芽伸长;pH 3.5处理抑制种子除发芽率外的其他萌发指标,也抑制根、芽长和生物量,特别是对根长和活力指数的抑制作用最大。何海洋等[17]研究发现pH 5.0~6.5对光皮桦种子萌发影响较小,且适当的弱酸处理会促进种子萌发。张玉秀等[18]研究表明,pH 3.0~5.0对紫穗槐种子的萌发影响较小,pH 4.5~5.0有利于其生长,pH < 4.5时胚根长、胚芽长和鲜重均逐渐减小,pH 3.0时根长和苗高会受到明显抑制,这与本试验的酸响应研究结果一致。
重金属在自然环境中很难被降解,土壤重金属污染对种子萌发和幼苗生长发育会产生很大的危害。Pb、Cd等重金属胁迫会使植物的生理生化发生一定程度的变化,抑制植物地上部、地下部的生长发育,使植物生物量、根长度和根体积等指标值减小[19]。本研究发现Cd 5处理下的种子发芽率、发芽势和发芽指数均受到促进,Cd 10则相反。陈俊任等[20]研究表明,低浓度的Cu和Zn对毛竹种子的发芽势、发芽率、发芽指数等指标有促进作用,高浓度则显著抑制,与本研究结果一致。另外,随着Cd浓度的增加,宽叶雀稗生物量逐渐减小,活力指数、根长、芽长均受到抑制,抑制作用表现为Cd 10>Cd 20>Cd 5,且在Cd 10和Cd 20下,根长的抑制作用大于芽长。张大鹏等[21]研究表明,Pb和Cd对毛竹种子的活力指数、幼苗根长、芽长及生物量等均具有显著的抑制作用,且抑制作用随着金属浓度的增大而增强,特别是根部。陈伟等[22]研究表明,4种草坪草种的活力指数均随着Cd浓度的增大而逐渐减小,且对胚根的抑制作用大于胚芽,重金属离子对种子活力和幼苗生长有较大影响。这与本实验的结果不尽相同,可能是因为宽叶雀稗在低浓度胁迫下,植物体内产生ROS的速度超过清除速度,造成ROS积累,对细胞造成损伤,但当胁迫浓度大于一定的阈值时,抗氧化酶和AsA-GSH循环等防御系统作用增强,清除ROS、超氧阴离子自由基、羟基自由基等的能力增强,一定程度上减弱了过氧化对植物的毒害[23-24]。
植物在遭受逆境后会产生大量ROS,使膜脂、蛋白质和生物大分子变性,膜透性增大,细胞膜脂质过氧化[25]。本研究中,根尖经染色后的着色程度不同,酸胁迫下颜色由浅到深为pH 5.5 < CK < pH 4.5 < pH 3.5,Cd胁迫下颜色由浅到深为CK < Cd 5 < Cd 20 < Cd 10,说明酸、Cd胁迫会影响根尖质膜的稳定性和功能性,根尖受到了不同程度的损伤,这与罗洁文等[26]Cd、Pb胁迫对类芦根系影响的研究结果一致。
植物的形态结构变化与机能的统一是植物适应生长环境的生物学基础[27]。研究发现,逆境胁迫会对植物细胞造成不同程度的损伤,一般表现为细胞壁、细胞膜被破坏,叶绿体、线粒体变形,被膜不完整,基粒弯曲、类囊体排列混乱和内嵴消失等现象[28]。本研究中,宽叶雀稗芽在CK、pH 5.5处理下,细胞结构完整,叶绿体、核糖体清晰可见,基粒和类囊体结构清晰,排列整齐。随着胁迫增强,在pH 3.5或Cd 10处理下,细胞结构遭到严重破坏,出现质壁分离、叶绿体破裂,质体小球变大、增多,甚至细胞空泡化现象。这与张黛静等[29]、田景花等[30]的研究结果一致。另外,本研究发现在Cd胁迫下,细胞壁、细胞器周围聚积的黑色颗粒物是重金属Cd。可能是宽叶雀稗细胞壁对Cd2+的固持作用有一定的阈值,细胞壁上的膜蛋白可以结合一定量的Cd2+来阻隔其进入细胞,但随着浓度的增大,膜结构遭到破坏,Cd2+进入细胞内对细胞器造成毒害,与黄玫英等[31]对Pb在类芦组织和亚细胞中的分布规律和毒害的研究结论一致。
4 结论(1)pH 3.5~5.5处理会促进宽叶雀稗种子发芽率,pH 4.5~6.1处理对发芽势、发芽指数、活力指数、根芽长和生物量影响较小,但pH 5.5处理对各萌发指标和芽长有促进作用,pH 3.5处理严重抑制宽叶雀稗种子萌发、幼苗生长和生物量。
(2)Cd 5、Cd 10、Cd 20处理对宽叶雀稗种子的发芽率、发芽势和发芽指数影响较小,但Cd 5处理对此3个指标有促进作用,同时其生物量也为Cd胁迫中最大;Cd胁迫对萌发指标、根长和芽长的抑制作用表现为Cd 10>Cd 20>Cd 5。
(3)pH 5.5~6.1处理对根尖质膜和芽亚细胞结构有轻微影响,pH 3.5和Cd 10处理严重损害根尖质膜,破坏芽亚细胞结构。
| [1] |
何芳, 徐友宁, 乔冈, 等. 中国矿山环境地质问题区域分布特征[J]. 中国地质, 2010, 37(5): 1520-1529. HE Fang, XU You-ning, QIAO Gang, et al. Regional distribution characteristics of mine environmental geological problems in China[J]. Geology in China, 2010, 37(5): 1520-1529. DOI:10.3969/j.issn.1000-3657.2010.05.028 |
| [2] |
胡远伟.重金属矿山废水的硫酸盐还原菌处理研究[D].昆明: 昆明理工大学, 2017. HU Yuan-wei. Study on the treatment of sulphate reducing bacteria in heavy metal mine wastewater[D]. Kunming: Kunming University of Science and Technology, 2017. |
| [3] |
周建军, 周桔, 冯仁国. 我国土壤重金属污染现状及治理战略[J]. 中国科学院院刊, 2014, 29(3): 315-320, 350. ZHOU Jian-jun, ZHOU Ju, FENG Ren-guo. Status of China's heavy metal contamination in soil and its remediation strategy[J]. Bulletin of Chinese Academy of Sciences, 2014, 29(3): 315-320, 350. |
| [4] |
王友生.稀土开采对红壤生态系统的影响及其废弃地植被恢复机理研究[D].福州: 福建农林大学, 2016. WANG You-sheng. Effect of rare earth mining on red soil ecosystem and revegetation mechanism in its wasteland[D]. Fuzhou: Fujian Agriculture and Forestry University, 2016. |
| [5] |
王友生, 侯晓龙, 吴鹏飞, 等. 长汀稀土矿废弃地土壤重金属污染特征及其评价[J]. 安全与环境学报, 2014, 14(4): 259-262. WANG You-sheng, HOU Xiao-long, WU Peng-fei, et al. Analysis of the characteristics and the evaluation of heavy metal pollutions in the deserted land-area left-over by the rare earth mining in Changting, Fujian[J]. Journal of Safety and Environment, 2014, 14(4): 259-262. |
| [6] |
王玉珍, 黄晓, 蔡丽平, 等. 不同温度条件下土壤黏着剂对宽叶雀稗种子发芽与幼苗生长的影响[J]. 草业学报, 2018, 27(4): 139-149. WANG Yu-zhen, HUANG Xiao, CAI Li-ping, et al. Effects of soil adhesive on seed germination and seedling growth of Paspalum wettseinii under different temperatures[J]. Acta Prataculturae Sinica, 2018, 27(4): 139-149. |
| [7] |
王友生, 吴鹏飞, 侯晓龙, 等. 稀土矿废弃地不同植被恢复模式对土壤肥力的影响[J]. 生态环境学报, 2015, 24(11): 1831-1836. WANG You-sheng, WU Peng-fei, HOU Xiao-long, et al. Effect of different revegetation model on soil properties in abandon mine area of rare earth[J]. Ecology and Environmental Sciences, 2015, 24(11): 1831-1836. |
| [8] |
侯晓龙.铅超富集植物金丝草对Pb胁迫的响应机制研究[D].福州: 福建农林大学, 2013. HOU Xiao-long. Response mechanism of Pb hyperaccumulator Pogonatherum crinitum to Pb stress[D]. Fuzhou: Fujian Agriculture and Forestry University, 2013. |
| [9] |
鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. LU Ru-kun. Methods of soil agricultural chemistry analysis[M]. Beijing: China Agricultural Science and Technology Press, 2000. |
| [10] |
Wang Y S, Yang Z M. Nitric oxide reduces aluminum toxicity by preventing oxidative stress in the roots of Cassia tora L[J]. Plant & Cell Physiology, 2006, 46(12): 1915-1923. |
| [11] |
唐琨, 朱伟文, 周文新, 等. 土壤pH对植物生长发育影响的研究进展[J]. 作物研究, 2013, 27(2): 207-212. TANG Kun, ZHU Wei-wen, ZHOU Wen-xin, et al. Research progress on effects of soil pH on plant growth and development[J]. Crop Research, 2013, 27(2): 207-212. DOI:10.3969/j.issn.1001-5280.2013.02.25 |
| [12] |
Siddique M, Siddique M T, Ali S, et al. Macronutrient assessment in apple growing region of Punjab[J]. Soil & Environment, 2009, 28(2): 184-192. |
| [13] |
王青青, 周强, 於丙军. 水培条件下不同pH值对香根草幼苗形态和生理特性的影响[J]. 植物资源与环境学报, 2014, 23(3): 59-64. WANG Qing-qing, ZHOU Qiang, YU Bing-jun. Effects of different pH values on seedling morphology and physiological characteristics of Vetiveria zizanioides under hydroponic culture[J]. Journal of Plant Resources and Environment, 2014, 23(3): 59-64. DOI:10.3969/j.issn.1674-7895.2014.03.08 |
| [14] |
徐开杰, 史丽丽, 王勇锋, 等. 水培条件下pH值对柳枝稷幼苗生长发育的影响[J]. 生态学报, 2015, 35(23): 7690-7698. XU Kai-jie, SHI Li-li, WANG Yong-feng, et al. Effect of the pH value on switchgrass seedling growth and development in hydroponics[J]. Acta Ecologica Sinica, 2015, 35(23): 7690-7698. |
| [15] |
Rajjou L, Duval M, Gallardo K, et al. Seed germination and vigor[J]. Annual Review of Plant Biology, 2012, 63(3): 507-533. |
| [16] |
Zhang H, Irving L J, McGill C, et al. The effects of salinity and osmotic stress on barley germination rate:Sodium as an osmotic regulator[J]. Annals of Botany, 2010, 106(6): 1027-1035. DOI:10.1093/aob/mcq204 |
| [17] |
何海洋, 胡春芹, 丁强强, 等. 不同pH对光皮桦种子萌发及幼苗生长的影响[J]. 西南林业大学学报, 2013, 33(5): 29-33, 39. HE Hai-yang, HU Chun-qin, DING Qiang-qiang, et al. Effect of pH value on seed germination and seedling growth of Betula luminifera[J]. Journal of Southwest Forestry University, 2013, 33(5): 29-33, 39. DOI:10.3969/j.issn.2095-1914.2013.05.005 |
| [18] |
张玉秀, 张倩, 李霞, 等. 不同pH与重金属Cd胁迫对紫穗槐萌发和幼苗生长的影响[J]. 环境化学, 2012, 31(10): 1569-1574. ZHANG Yu-xiu, ZHANG Qian, LI Xia, et al. Effects of pH and cadmium on the seed germination and seedling growth of Amorpha fruticosa L[J]. Environmental Chemistry, 2012, 31(10): 1569-1574. |
| [19] |
陶玲, 任珺, 祝广华, 等. 重金属对植物种子萌发的影响研究进展[J]. 农业环境科学学报, 2007, 26(增刊1): 52-57. TAO Ling, REN Jun, ZHU Guang-hua, et al. Advance on the effects of heavy metals on seed germination[J]. Journal of Agro-Environment Science, 2007, 26(Suppl1): 52-57. |
| [20] |
陈俊任, 柳丹, 吴家森, 等. 重金属胁迫对毛竹种子萌发及其富集效应的影响[J]. 生态学报, 2014, 34(22): 6501-6509. CHEN Jun-ren, LIU Dan, WU Jia-sen, et al. Seed germination and metal accumulation of moso bamboo (Phyllostachy pubescens) under heavy metal exposure[J]. Acta Ecologica Sinica, 2014, 34(22): 6501-6509. |
| [21] |
张大鹏, 蔡春菊, 范少辉, 等. 重金属Pb2+和Cd2+对毛竹种子萌发及幼苗早期生长的影响[J]. 林业科学研究, 2012, 25(4): 500-504. ZHANG Da-peng, CAI Chun-ju, FAN Shao-hui, et al. Effects of Pb2+, Cd2+ on germination and seedling early growth of moso bamboo (Phyllostachys edulis) seed[J]. Forest Research, 2012, 25(4): 500-504. DOI:10.3969/j.issn.1001-1498.2012.04.014 |
| [22] |
陈伟, 张苗苗, 宋阳阳, 等. 重金属胁迫对4种草坪草种子萌发的影响[J]. 草地学报, 2013, 21(3): 556-563. CHEN Wei, ZHANG Miao-miao, SONG Yang-yang, et al. Inhibitory effect of heavy metal stress on the seed germination of four turfgrass types[J]. Acta Agrestia Sinica, 2013, 21(3): 556-563. |
| [23] |
韩航, 陈顺钰, 赵雅曼, 等. 铅胁迫对金丝草AsA-GSH循环及铅积累的影响[J]. 农业环境科学学报, 2018, 37(4): 656-664. HAN Hang, CHEN Shun-yu, ZHAO Ya-man, et al. Influence of lead stress on the ascorbate-glutathione cycle and subcellular distribution in leaves and roots of Pogonatherum crinitum[J]. Journal of Agro-Environment Science, 2018, 37(4): 656-664. |
| [24] |
何洁, 高钰婷, 贺鑫, 等. 重金属Zn和Cd对翅碱蓬生长及抗氧化酶系统的影响[J]. 环境科学学报, 2013, 33(1): 312-320. HE Jie, GAO Yu-ting, HE Xin, et al. The effect of Zn and Cd on growth and antioxidant enzymes activity of Suaeda heteroptera Kitagawa[J]. Acta Scientiae Circumstantiae, 2013, 33(1): 312-320. |
| [25] |
贾学静, 董立花, 丁春邦, 等. 干旱胁迫对金心吊兰叶片活性氧及其清除系统的影响[J]. 草业学报, 2013, 22(5): 248-255. JIA Xue -jing, DONG Li-hua, DING Chun-bang, et al. Effects of drought stress on reactive oxygen species and their scavenging systems in Chlorophytum capense var.medio-pictum leaf[J]. Acta Prataculturae Sinica, 2013, 22(5): 248-255. |
| [26] |
罗洁文, 李莹, 苏烁烁, 等. 类芦根系抗氧化酶和植物螯合肽对Cd、Pb胁迫的应答[J]. 生态环境学报, 2016, 25(6): 1047-1053. LUO Jie-wen, LI Ying, SU Shuo-shuo, et al. Response of antioxidant enzymes and PCs in root of Neyraudiareynaudiana to Cd, Pb stress[J]. Ecology and Environmental Sciences, 2016, 25(6): 1047-1053. |
| [27] |
刘冉, 石峰, 刘伟成, 等. 不同形态氮素对盐胁迫下番茄细胞超微结构与光合作用的影响[J]. 园艺学报, 2015, 42(3): 471-479. LIU Ran, SHI Feng, LIU Wei-cheng, et al. Effect of nitrogen forms on cell ultrastructure and photosynthesis of tomato under salinity[J]. Acta Horticulturae Sinica, 2015, 42(3): 471-479. |
| [28] |
徐萍, 李进, 吕海英, 等. 干旱胁迫对银沙槐幼苗叶绿体和线粒体超微结构及膜脂过氧化的影响[J]. 干旱区研究, 2016, 33(1): 120-130. XU Ping, LI Jin, LÜ Hai-ying, et al. Effects of drought stress on ultrastructure of chloroplast and mitochondria and membrane lipid peroxidation of Ammodendron argenteum[J]. Arid Zone Research, 2016, 33(1): 120-130. |
| [29] |
张黛静, 马建辉, 杨淑芳, 等. 硅对铜胁迫下小麦幼根细胞超微结构的影响[J]. 应用生态学报, 2014, 25(8): 2385-2389. ZHANG Dai-jing, MA Jian-hui, YANG Shu-fang, et al. Effects of silicon on the ultrastructures of wheat radical cells under copper stress[J]. Chinese Journal of Applied Ecology, 2014, 25(8): 2385-2389. |
| [30] |
田景花, 王红霞, 张志华, 等. 低温逆境对不同核桃品种抗氧化系统及超微结构的影响[J]. 应用生态学报, 2015, 26(5): 1320-1326. TIAN Jing-hua, WANG Hong-xia, ZHANG Zhi-hua, et al. Effects of chilling stress on antioxidant system and ultrastructure of walnut cultivars[J]. Chinese Journal of Applied Ecology, 2015, 26(5): 1320-1326. |
| [31] |
黄玫英, 罗洁文, 黄彩凤, 等. Pb在类芦组织和亚细胞中的分布规律和毒害效应[J]. 农业环境科学学报, 2016, 35(11): 2077-2085. HUANG Mei-ying, LUO Jie-wen, HUANG Cai-feng, et al. The distribution and toxic effects of Pb at the levels of the tissue and sub-cellular in Neyraudia reynaudiana[J]. Journal of Agro-Environment Science, 2016, 35(11): 2077-2085. DOI:10.11654/jaes.2016-0627 |