 2019, Vol. 38
2019, Vol. 38



随着国防军工和核电工业对铀资源的需求日益增大,在开采和利用铀资源的过程中不可避免地要产生大量的铀矿冶废石和铀尾矿,其中一定数量的铀[1],经过雨水和地下水的作用会进入周边环境并可能通过食物链进入人体,以内照射和化学毒性的方式损害人体健康[2]。因此,针对铀污染的环境修复成为国内外广大学者研究的重点。
针对铀污染土壤修复的方法有物理法、化学法和生物修复法。物理法和化学法由于工程量较大、修复成本高等因素很难推广,而生物修复法因为其不会破坏土壤结构,不易对土壤造成二次污染而得到学者的广泛认同。植物修复技术是指运用植物遏制、降解或提取水中或土壤基质中的外源性物质的技术[3]。与其他生物修复技术相比,植物修复具有环保、绿色、无公害、成本低的优点。
黑麦草(Lolium perenne L.)为草本植物,生长速度快,适应性广,是引种栽培的优良牧草,也常常作为水土保持的先锋植物,在草原建设及畜牧业发展中具有重要作用,近几年发现其对重金属如铜、镉、铅、汞等也有较强的富集能力[4-7]。针对植物修复技术的大多数研究主要集中于寻找理想的超富集植物、了解植物在铀污染土壤环境中的适应能力和探究铀胁迫下植物生长的机制。关于不同浓度铀污染土壤中作物体内各部位铀积累分布特征及动态变化的研究极少。因此,本文以盆栽试验的方式,研究在不同浓度铀污染土壤下,铀对黑麦草光合参数短期内的动态变化、土壤铀残留浓度和黑麦草植株体各部分的短期积累的动态变化,为植物修复重金属或放射性核素污染土壤提供一定的理论基础。
1 材料与方法 1.1 供试植物与土壤试验选择禾本科的黑麦草(Lolium perenne L.)作为供试植物。试验场地位于西南科技大学材料环境降解及安全性评价试验场,选用壤土,采自西南科技大学农学试验基地菜园,土壤理化性质为:有机质19.24 g · kg-1,总N、P、K分别为1.23、0.43 g · kg-1和12.18 g·kg-1,速效N、P、K分别为136.3、7.8 mg·kg-1和108 mg·kg-1,土壤pH 4.51,土壤中铀含量为3.5~4.0 mg·kg-1。
1.2 试验设计与处理方法试验设计:土壤处理采取向土壤中添加外源铀污染的方法,所用药品为乙酸双氧铀,试验设置4个外源铀浓度:25、50、75、100 mg·kg-1,同时设置空白对照CK,共取3周,每处理设置3个重复,共45盆。盆规格为Φ16 cm×13 cm,每盆装干质量1 kg土壤。处理方法:于2018年3月用饱和持水量法进行土壤铀污染处理,以乙酸双氧铀的形式按每盆350 mL的量配成相应浓度的溶液施入土壤中,空白组施加等量清水。土壤处理后用黑色遮阳网覆盖放置于通风干燥处。铀处理土壤稳定8周以后再栽种植物,保证铀被土壤充分吸附。每盆栽种5株,保持田间持水量的60%~ 70%,黑麦草生长56、63 d和70 d后测定叶绿素荧光参数、光合参数,并分别采集土壤样品和植物样品。
1.3 叶绿素荧光参数测定叶绿素荧光参数测定采用M-PEA-2荧光仪(英国Hansatech公司),测量前先将叶片暗适应20 min,再进行测定[8]。测定了反映PSⅡ反应中心内禀光能转换效率的最大光化学效率(Fv/Fm)、反映J点的相对可变荧光强度的相对可变荧光(Vj)和以吸收光能为基础的性能指数——光合性能指数(PIabs)。
1.4 光合气体交换参数的测定采用LC Pro-SD+全自动便携式光合仪(澳作生态仪器有限公司),在自然光照、大气CO2浓度、温度和湿度下,分别于56、63 d和70 d上午9:00—11:30测定黑麦草叶片光合气体交换参数:光合速率(A)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。每个处理测3次,取平均值[9]。
1.5 土壤铀含量测定土壤样品放入80 ℃烘箱中烘至恒质量,使用中药粉碎机(北京永光明)粉碎,称取0.200 0 g,加入5 mL浓硝酸、2 mL氢氟酸、2 mL盐酸和2 mL 30%双氧水,使用Synthos 3000消解仪(奥地利安东帕公司)消解,消解液通过滤膜过滤后使用Agilent 7700x ICPMS(美国安捷伦公司)测定铀含量。
1.6 植物各部位铀富集量的测定植物样品用自来水洗净,再用超纯水清洗3次,滤纸擦干,放入105 ℃烘箱中杀青30 min,而后80 ℃烘至恒质量。使用中药粉碎机(北京永光明)粉碎,称取0.200 g,加入7 mL浓硝酸和2 mL 30%双氧水,使用Synthos 3000消解仪(奥地利安东帕公司)消解,消解液通过滤膜过滤后使用Agilent 7700x ICP-MS(美国安捷伦公司)测定铀含量。
1.7 计算方法与数据分析生物富集系数(BCF)=植物干物质中重金属含量(mg·kg-1 DW)/土壤重金属浓度(mg·kg-1 DW)[9]。
生物转移因子(TF)=植物地上部干物质重金属浓度(mg·kg-1 DW)/植物地下部干物质重金属浓度(mg·kg-1 DW)[9]。
铀总提取量=植物地下部干物质中铀浓度×地下部干质量+植物地上部干物质铀浓度×地上部干质量。
使用WPS 2018进行数据的整理,使用DPS7.05[10]进行方差分析,分析方法为Tukey多重比较法,P < 0.05为显著,P < 0.01为极显著,应用Origin 8软件作图,并用其分析功能进行线性拟合和非线性拟合。
2 结果与分析 2.1 黑麦草对铀的富集特性由图 1(A)可以看出,以黑麦草作为修复植物,对外源铀污染土壤进行修复后,土壤的铀残留浓度随外源铀浓度的增高而增高(P < 0.05),在56 d 100 mg·kg-1处理下达到最大,为96.46 mg·kg-1,然而在63 d和70 d 100 mg·kg-1时,土壤的铀残留浓度为73.06 mg·kg-1和62.53 mg·kg-1( P < 0.05),说明黑麦草对不同浓度铀污染的土壤是有修复效果的。其中,100 mg·kg-1外源铀处理下,土壤铀残留浓度减少值最大,分别比56 d减少了23.39 mg·kg-1和33.92 mg·kg-1,而25 mg·kg-1时,土壤铀浓度减少值最小,分别比56 d减少了7.59 mg·kg-1和8.99 mg·kg-1。图 1(B)和图 2(A)分别是铀处理浓度和周数与土壤实际铀浓度之间的关系,对其拟合,得知外源铀浓度与土壤实际浓度呈线性关系,且斜率随着生长时间的增加而减小,而决定系数(R2)为0.99、0.98、0.98,说明外源铀浓度对土壤铀残留浓度的影响较大;生长时间与土壤实际铀浓度呈指数衰减的趋势,最后逐渐趋于稳定。

|
图中不同小写字母表示P < 0.05水平上的差异显著。下同 Different lowercase letters in the figure indicate significant differences at P < 0.05 level. The same below 图 1 不同生长时间下土壤(A、B)、地下部(C、D)、地上部(E、F)铀浓度与外源铀处理浓度的关系 Figure 1 Relationship between uranium concentration in soil(A, B), underground(C, D)and aboveground(E, F)and treatment concentration of exogenous uranium under different growth time |

|
图 2 不同外源铀浓度下土壤(A)、地下部(B)、地上部(C)铀浓度与生长时间的关系 Figure 2 Relationship between uranium concentration and time in soil(A), underground(B)and aboveground(C)parts under different U concentrations |
由图 1(C)可知在56 d时,不同浓度铀污染的土壤中,黑麦草地下部的铀浓度分别为3.29、96.09、185.15、79.05、137.20 mg·kg-1,在63 d时,黑麦草地下部的铀浓度达到了3.31、131.96、308.73、442.22、905.40 mg · kg-1,在56 d的基础上增长了0.62%、37.33%、66.75%、459.43%、559.50%,在70 d时,黑麦草地下部的铀浓度分别为4.71、193.55、375.03、534.53 mg·kg-1和1070.63 mg·kg-1,在56 d的基础上增长了43.20%、101.42%、102.56%、576.20%、680.33%。说明不同浓度污染下黑麦草地下部铀浓度随生长时间的增加而增加(P < 0.05)。根据图 1(D)可知地下部铀浓度与外源铀处理浓度呈线性关系,在70 d的决定系数(R2)较高,为0.90,说明随着生长时间的增加外源铀处理浓度对黑麦草地上部的铀浓度影响逐渐密切。根据图 2(B)的非线性拟合可知,在高浓度处理下,56~63 d黑麦草地下部铀浓度的增长量比63~70 d之间的增长量大,这种差异在100 mg·kg-1处理下为最大。图 2(A)和图 2(B)在56~63 d的时候土壤中的铀含量减少较多,而黑麦草地下部含量在56~63 d增加的较多,说明在56~63 d这个生长时间段内黑麦草地下部富集铀的能力较强,而63~70 d铀浓度的浓度差较小的原因可能是因为根部富集的铀逐渐达到峰值,因此63~70 d土壤的铀实际浓度差的也较小。
由图 1(E)可知在56 d时,不同浓度铀污染的土壤中,黑麦草地上部的铀浓度分别为1.53、23.62、88.85、95.04、237.34 mg·kg-1,即随外源铀浓度的增加而增加(P < 0.05),在63 d时,黑麦草地上部的铀浓度达到了3.42、116.46、135.93、179.06、340.30 mg·kg-1,在56 d的基础上增长了123.51%、392.96%、52.99%、88.40%、43.38%,在70 d时,黑麦草地上部的铀浓度分别为2.65、151.59、250.44、351.71 mg · kg-1和628.33 mg · kg-1,在56 d的基础上增长了73.07%、541.79%、181.87%、270.07%、167.74%。说明不同浓度污染下黑麦草地上部铀浓度随生长时间的增加而增加(P < 0.05)。斜率随生长时间的增大而增大,说明地上部铀浓度与外源铀浓度正相关,且随着生长时间的增加铀处理浓度对黑麦草地上部的铀浓度影响也越来越密切。而根据图 2(B)和图 2(C)与生长时间的非线性拟合可知在中高浓度铀处理下黑麦草地上部铀浓度在63~70 d时的浓度差大于56~63 d的浓度差,而黑麦草地下部在56~63 d时的浓度差比63~70 d高,说明在56~63 d时黑麦草地上部虽然会富集一定量的铀,但其主要富集在地下部,但在63 d后,虽然铀还是主要富集在地下部,但是铀逐渐在向地上部转移,且这种转移趋势较快。
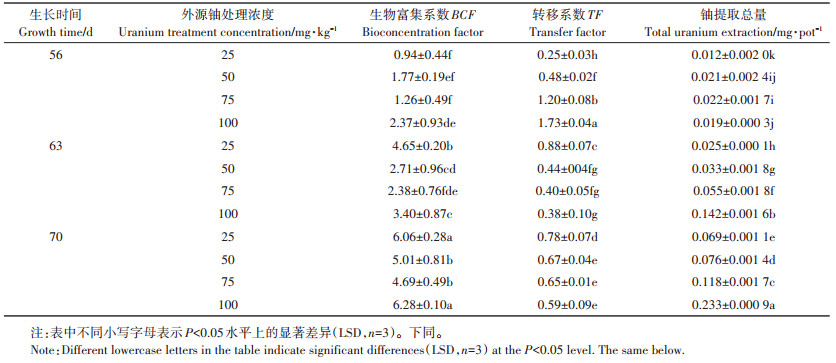
由表 1可知, 当外源铀浓度为0~100 mg·kg-1时,黑麦草在不同生长时间下的转移系数(TF)均逐渐增加,但生物富集系数(BCF)却呈下降趋势,分别为0.94~6.06(25 mg·kg-1)、1.77~5.01(50 mg·kg-1)、1.26~ 4.69(75 mg·kg-1)和2.37~6.28(100 mg·kg-1)。当外源铀为100 mg·kg-1时,黑麦草的铀提取总量在56、63、70 d时分别达到0.019、0.142、0.233 mg,呈明显的“累积效应”。同时,随黑麦草生长时间的延长,BCF也在逐渐增加。这表明,黑麦草根系对土壤铀具有较强的吸收能力,且黑麦草对土壤铀的累积、转移特征与外源铀污染水平、生长时间密切相关。
|
|
表 1 不同浓度铀处理下BCF、TF、铀提取总量随生长时间的动态变化 Table 1 Dynamic change of BCF, TF, total uranium extraction with time under different U treatments |
由图 3可以看出,随着外源铀浓度的增加,不同时期黑麦草的光合性能指数(PIabs)呈现先上升后下降的趋势(P < 0.05),而随着生长时间的增加,不同浓度处理下的黑麦草PIabs也呈现先上升后下降的趋势(P < 0.05),在56 d时,与对照组相比黑麦草的PIabs变化了-2.10%、46.96%、16.23%、17.67%;在70 d时,与对照组相比黑麦草的PIabs变化了7.09%、10.47%、-0.42%、-26.75%。

|
图 3 不同浓度铀处理下黑麦草光合性能指数PIabs随生长时间的动态变化 Figure 3 Dynamic changes of PIabs with ryegrass under different concentrations of U treatment |
由图 4可以看出,最大光化学效率(Fv/Fm)随外源铀浓度的增加和植物生长时间的增加,呈现较为平稳的趋势,总体是维持在0.771~0.804的区间范围内,外源铀浓度和生长时间对黑麦草的最大光化学效率影响不显著。

|
图 4 不同浓度铀处理下黑麦草Fv/Fm随生长时间的动态变化 Figure 4 Dynamic changes of Fv/Fm with ryegrass under different concentrations of U treatment |
由图 5可知,不同生长时间的植物叶片相对可变荧光(Vj)随铀浓度的升高呈现先增高后降低的趋势,在70 d 25 mg·kg-1外源铀浓度处理下达到最大,为0.48,比56 d 25 mg·kg-1处理的Vj增加了7.09%,比70 d CK组增加了13.66%。

|
图 5 不同浓度铀处理下黑麦草Vj随生长时间的动态变化 Figure 5 Dynamic changes of Vj with ryegrass under different concentrations of U treatment |
有文献指出外源重金属胁迫不仅对植物的光合荧光参数产生影响,也会对植物的光合气体参数产生影响,针对外源铀污染胁迫,本研究对黑麦草光合气体参数进行了检测。
由表 2可知,在不同的外源铀浓度和生长时间的处理下,黑麦草的PSⅠ的几个指标呈现出不同的规律。黑麦草的光合速率(A)和气孔导度(Gs)在不同生长时间和不同浓度铀胁迫下都小于CK,但是随外源铀浓度的变化和生长时间变化的差异不显著。胞间CO2浓度(Ci)在不同生长时间和不同浓度处理下的值相比差异不显著,其中在70 d 75 mg·kg-1时达到最大,分别比70 d CK和56 d 75 mg·kg-1增加了35.19%和38.86%。蒸腾速率(Tr)是在56 d 75 mg·kg-1时达到最大,比56 d CK增加了14.73%,且蒸腾速率的值维持在3.50~6.62。
|
|
表 2 黑麦草光合作用参数对土壤铀污染的生理响应 Table 2 Dynamic changes of biological effect index(BI)of photosynthetic system Ⅰ (PSⅠ)of plants under different concentrations of U-contaminated soil |

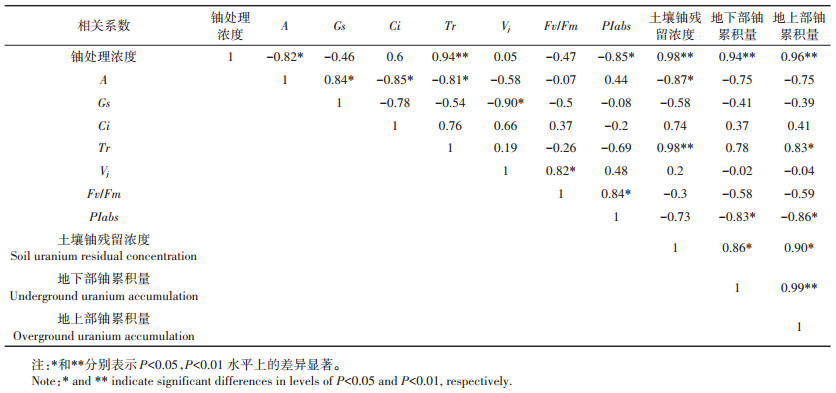
将不同生长时间下的光合速率(A)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、相对可变荧光(Vj)、光合性能指数(PIabs)、最大光化学效率(Fv/ Fm)、土壤铀残留浓度、地下部铀累计量和地上部铀累计量计算平均值,然后分析其相关性。
由表 3可知,外源铀浓度与黑麦草的Tr、土壤铀残留浓度、地下部铀累积量和地上部铀累积量极显著相关;黑麦草的Tr与土壤铀残留浓度极显著相关,与地上部累积量显著相关;PIabs与Fv/Fm、地下部铀累积量、地上部铀累积量显著相关;土壤铀残留浓度与地下部铀累积量、地上部铀累积量显著相关;地下部铀累积量与地上部铀累积量极显著相关。
|
|
表 3 光合特性与黑麦草富集量之间的相关性分析 Table 3 Correlation analysis between photosynthetic characteristics and ryegrass enrichment |
本研究中,黑麦草植物体内铀浓度随土壤铀残留浓度升高而升高,且地上部和地下部与铀处理浓度呈正相关,随着生长时间的增加,外源铀浓度对黑麦草地上部和地下部的铀浓度影响逐渐明显,说明影响黑麦草体内的铀浓度主要与外源铀污染浓度有关,这与徐国聪等[11]的研究结果基本一致。各处理地下部铀浓度均高于地上部,且地下部和地上部铀浓度随生长时间的增加呈指数增加的形式,且地上部增加的幅度要比地下部高,说明随着单位时间的增加,黑麦草地下部富集铀的浓度逐渐降低,并向地上部运输,使得地上部的单位时间内的铀浓度逐渐增大,这与廖若星等[12]的研究结果相似。
富集系数[13](BCF)是指植株体内元素含量与环境元素含量之比,其值越大,则表明植物吸收能力越强。转运系数[13](TF)是指地上部分铀的含量与地下部分铀的含量之比,它表示铀元素在植株体内转运情况,可作为评价植物将铀从地下部分向地上部分转运能力大小的指标。BCF和TF分别在70 d 100 mg·kg-1和56 d 100 mg·kg-1处理时达到最大值,这是因为在高水平土壤铀浓度下,土壤中铀含量基数较大。同时也表明,随着生长时间的增加,黑麦草将铀主要富集在地下部。多数学者认为[14-16],重金属超富集植物应满足3个指标:植株体重金属浓度达到相应标准(Cd为100 mg·kg-1,Pb为1000 mg·kg-1,Zn、Mn为10 000 mg·kg-1,等);BCF>1;TF>1。按照这一标准,在本文试验条件下黑麦草满足BCF>1这一条件,但其TF在本研究中多数都小于1,因此黑麦草不是铀的超富集植物。传统超富集植物评价指标最大的不足是忽略了对植物生物量的讨论,因此有学者[17-20]提出了生物富集量系数、转运量系数、单株植物总提取(剔除)量、吸收模量、修复年限、修复指数量等指标,引入了生物量及植物生长周期对修复效果的考虑。黑麦草具有生物量大、生长周期短、再生能力强、种植管理方便、对铀胁迫耐受性强等优点,有望通过合理的农艺管理措施,增加其对铀的绝对富集量,强化其修复能力。
叶绿素荧光参数的变化能够反映外界的环境胁迫对植物光合作用各过程产生的影响,因此将叶绿素荧光参数作为光合反应中的灵敏指标[21]。Fv/Fm[22]的值反映了PSⅡ反应中心均处于开放状态时的量子产量,即PSⅡ的潜在最大量子产量。在本试验中不同外源铀浓度处理下,黑麦草的Fv/Fm与对照都在正常范围内,并无明显的差异。PIabs[23]表示光合作用性能指数,此性能指数可以更准确地反映生物PSⅡ的生理活性状态,能够较准确合理地反映胁迫对光系统的影响[23-24]。本研究结果表明,在铀胁迫下,黑麦草的PIabs呈先增后减的趋势,说明黑麦草在中低浓度下PSⅡ耐受性较好,Vj [23]表示叶绿素荧光动力学曲线中J点的相对可变荧光,本试验中黑麦草的Vj的呈先升后降的趋势,但总体上维持在0.4~0.5。Fv/Fm、PIabs和Vj能够反映外界的胁迫对于光合系统Ⅱ的影响,在外源铀胁迫下Fv/Fm、Vj没有显出显著性差异,而PIabs呈先增后减的趋势,说明黑麦草的光合系统Ⅱ整体上对外源铀胁迫的抗性较高,并未对光合系统Ⅱ产生影响。
重金属胁迫会对植物的光合作用过程造成影响,最终导致碳固定和同化效率降低[25-26]。有研究表明[27-29],当A下降时,若植物Ci增加,说明引起叶片光合速率降低的原因是叶肉细胞光合活性下降,即非气孔因素;若Ci减小,则为气孔因素。本研究显示,黑麦草在铀胁迫下,其光合速率(A)、气孔导度(Gs)显著降低,胞间CO2浓度(Ci)的改变则不显著,因此不能判断黑麦草光合速率(A)下降是由非气孔因素还是气孔因素引起的。也有研究[30-31]表明光合速率下降的内在原因可能因光抑制引起的,即光合机构吸收的光能超过光合作用本身所能利用的能量而引起光合效率下降。本次试验的植物生长环境都有遮阴和通风扇的大棚,在测量时,外遮和内遮都会打开并关闭风扇,使用日光测植物的光合参数,所以猜测植物在测量时受到了光抑制。
4 结论各浓度铀处理对植物黑麦草的光合气体交换参数尤其是光合速率会有显著影响,可能是非气孔因素导致的,而不同生长时间段各浓度处理对黑麦草光合荧光参数影响不显著,对光合系统PSⅡ影响较小。外源铀污染浓度对黑麦草地下部和地上部铀浓度随生长时间的增加,影响逐渐增加,单位时间内的铀浓度变化地下部小于地上部,但地下部富集量大于地上部。本试验中黑麦草虽未达到铀超富集植物的指标,但其较强的耐受性和较高的富集量使其成为铀污染土壤植物修复的潜在植物。
| [1] |
任德曦, 胡泊. 论推进我国完整的核能产业经济体系的发展[J]. 南华大学学报(社会科学版), 2014, 1502: 1-9. REN De-xi, HU Bo. On promoting the development of China's complete nuclear energy industry economic system[J]. Journal of Nanhua University (Social Science Edition), 2014, 1502: 1-9. |
| [2] |
杨巍, 杨亚新, 曹龙生, 等. 某铀尾矿库中放射性核素对环境的影响[J]. 东华理工大学学报(自然科学版), 2011, 3402: 155-159. YANG Wei, YANG Ya-xin, CAO Long-sheng, et al. Effects of radionuclides on the environment in a uranium tailings reservoir[J]. Journal of East China Institute of Technology(Natural Science Edition), 2011, 3402: 155-159. |
| [3] |
Chang P, Kim K. Uranium accumulation of crop plants enhanced by citric acid[J]. Environmental Geochemistry and Health, 2005, 27(5/6): 529-538. |
| [4] |
张刚, 翁悦, 李德香, 等. 铜胁迫对黑麦草种子萌发及幼苗生理生态的影响[J]. 东北师大学报(自然科学版), 2019, 51(1): 119-124. ZHANG Gang, WENG Yue, LI De-xiang, et al. Growth and physiology responses of perennial ryegrass to Cu2+ stress[J]. Journal of Northeast Normal University (Natural Science Edition), 2019, 51(1): 119-124. |
| [5] |
马婵华. 黑麦草植物对农田重金属镉污染土壤的修复效果研究[J]. 现代农业科技, 2019(3): 148-152. MA Chan-hua. Study on rehabilitation effect of ryegrass plants on heavy metal cadmium contaminated soil in farmland[J]. Modern Agricultural Science and Technology, 2019(3): 148-152. |
| [6] |
李瑞娟, 董雨滔, 张帅. 黑麦草修复土壤铅污染研究进展[J]. 农业工程, 2019, 9(1): 44-46. LI Rui-juan, DONG Yu-tao, ZHANG Shuai. Review on remediation of lead pollution in soil by ryegrass[J]. Agricultural Engineering, 2019, 9(1): 44-46. |
| [7] |
过昱辰.三种草坪草对土壤汞污染耐受性及富集效果的研究[D].苏州: 苏州大学, 2016. GUO Yu-chen. Study on tolerance and enrichment effect of three kinds of turf grass on soil mercury pollution[D]. Suzhou: Suzhou University, 2016. |
| [8] |
Schansker G, Srivastava A, Strasser R J. Characterization of the 820-nm transmission signal paralleling the chlorophyll afluorescence rise (OJIP)in pea leaves[J]. Functional Plant Biology, 2003, 30(7): 785-796. |
| [9] |
贾文甫, 罗学刚, 陈功亮, 等. 黄秋葵对铀胁迫的光合生理响应及吸收特征[J]. 环境科学与技术, 2015, 38(10): 41-47. JIA Wen-fu, LUO Xue-gang, CHEN Gong-liang, et al. photosynthetic physiological response and absorption characteristics of okra under uranium stress[J]. Environmental Science and Technology, 2015, 38(10): 41-47. |
| [10] |
唐启义, 冯明光. DPS数据处理系统——实验设计统计分析及数据挖掘[M]. 北京: 科学出版社, 2010. TANG Qi-yi, FENG Ming-guang. DPS data processing system:Experimental design, statistical analysis and data mining[M]. Beijing: Science Press, 2010. |
| [11] |
徐国聪, 唐运来, 陈梅, 等. 铀对菠菜叶片光合作用影响的研究[J]. 西北植物学报, 2016, 36(2): 370-376. XU Guo-cong, TANG yun-lai, CHEN mei, et al. Study on the effect of uranium on photosynthesis of spinach leaves[J]. Northwest Plant Journal, 2016, 36(2): 370-376. |
| [12] |
廖若星, 赖金龙, 李月琴, 等. 蚕豆对土壤中锶的累积及其器官分配特征分析[J]. 农业环境科学学报, 2017, 36(10): 1953-1959. LIAO Ruo-xing, LAI Jin-long, LI Yue-qin, et al. Analysis of the accumulation of earthworm and its organ allocation in broad bean[J]. Environmental Science & Technology, 2017, 36(10): 1953-1959. |
| [13] |
王帅.某铀矿区农田土壤重金属形态分析及其风险评价[D].南昌: 东华理工大学, 2017. WANG Shuai. Speciation analysis and risk assessment of heavy metals in farmland soils in uranium mining areas[D]. Nanchang: East China University of Technology, 2017. |
| [14] |
万芹方, 陈雅宏. 植物对土壤中铀的吸收与富集[J]. 植物学报, 2011, 46(4): 425-436. WAN Qin-fang, CHEN Ya-hong. Uptake and enrichment of uranium in soil by plants[J]. Chinese Journal of Plant Science, 2011, 46(4): 425-436. |
| [15] |
Baker A J M, Brooks R R. Terrestrial higher plants of hypercumulation metallic element:A review of their distribution, ecology and phytochemistry[J]. Biorecovery, 1989, 1(2): 81-126. |
| [16] |
沈振国, 刘友良. 重金属超积累植物的研究进展[J]. 植物生理学通讯, 1998, 34(2): 133-139. SHEN Zhen-guo, LIU You-liang. Research progress of heavy metal hyperaccumulators[J]. Plant Physiology Communications, 1998, 34(2): 133-139. |
| [17] |
唐永金, 罗学刚. 植物吸收和富集核素的研究方法[J]. 核农学报, 2011, 25(6): 1292-1299. TANG Yong-jin, LUO Xue-gang. Research methods for plant absorption and enrichment of nuclides[J]. Journal of Nuclear Agricultural Sciences, 2011, 25(6): 1292-1299. |
| [18] |
聂发辉. 关于超富集植物的新理解[J]. 生态环境, 2005, 14(1): 136-138. NIE Fa-hui. A new understanding of hyperaccumulators[J]. Ecological Environment, 2005, 14(1): 136-138. |
| [19] |
Wang H X, Shi G X. Photosynthetic physiological characteristics and ultrastructure of Salvinia natans leaves under Cr6+ stress[J]. Acta Bot Boreal Occident Sin, 2008, 28(11): 2244-2250. |
| [20] |
郑九华, 冯永军, 于开芹, 等.重金属污染土壤的植物修复评价指标的理论探讨[C].第二届重金属污染监测风险评价及修复技术高级研讨会, 2008: 220-225. ZHENG Jiu-hua, FENG Yong-jun, YU Kai-qin, et al. Theoretical discussion on evaluation index of phytoremediation of heavy metal contaminated soil[C]. The 2nd Seminar on Risk Assessment and Remediation Technology for Heavy Metal Pollution Monitoring, 2008: 220-225. |
| [21] |
Peter J R, Rolf G. Rapid light curves:A powerful tool to assess photosynthetic activity[J]. Aquatic Botany, 2005, 82(3): 222-237. |
| [22] |
汪月霞, 孙国荣, 王建波, 等. NaCl胁迫下星星草幼苗MDA含量与膜透性及叶绿素荧光参数之间的关系[J]. 生态学报, 2006(1): 122-129. WANG Yue-xia, SUN Guo-rong, WANG Jian-bo, et al. Relationship between MDA content and membrane permeability and chlorophyll fluorescence parameters of Puccinellia tenuiflora seedlings under NaCl stress[J]. Acta Ecologica Sinica, 2006(1): 122-129. |
| [23] |
李鹏民, 高辉远, Reto J S. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J]. 植物生理与分子生物学学报, 2005(6): 559-566. LI Peng-min, GAO Hui-yuan, Reto J S. Application of rapid chlorophyll fluorescence induction kinetic analysis in photosynthesis research[J]. Journal of Plant Physiology and Molecular Biology, 2005(6): 559-566. |
| [24] |
张朝轩, 杨天仪, 吴淑杭, 等. 微生物肥料对土壤生态及葡萄叶片叶绿素荧光特性的影响[J]. 天津农业科学, 2011, 17(1): 92-95. ZHANG Chao-xuan, YANG Tian-yi, WU Shu-hang, et al. Effects of microbial fertilizers on soil ecology and chlorophyll fluorescence characteristics of grape leaves[J]. Tianjin Agricultural Sciences, 2011, 17(1): 92-95. |
| [25] |
刘飞虎. 植物品种净光合速率测定的取样技术——以苎麻为例[J]. 云南大学学报(自然科学版), 2010, 32(2): 221-226. LIU Fei -hu. Sampling technique for determination of net photosynthetic rate of plant varieties:Taking castor as an example[J]. Journal of Yunnan University(Natural Science Edition), 2010, 32(2): 221-226. |
| [26] |
曹玲, 王庆成, 崔东海. 土壤镉污染对四种阔叶树苗木叶绿素荧光特性和生长的影响[J]. 应用生态学报, 2006(5): 769-772. CAO Ling, WANG Qing-cheng, CUI Dong-hai. Effects of soil cadmium pollution on chlorophyll fluorescence characteristics and growth of four broad-leaved tree seedlings[J]. Journal of Applied Ecology, 2006(5): 769-772. |
| [27] |
许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯, 1997(4): 241-244. XU Da-quan. Photosynthesis, stomatal limit the number of issues analysis[J]. Plant Physiology Communications, 1997(4): 241-244. |
| [28] |
Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology, 1982(33): 317-345. |
| [29] |
Bragina T V, Drozdova I S, Ponomareva Y V, et al. Photo-synthesis, respiration and transpiration in maize seedlings under hypoxia induced by complete flooding[J]. Doklady Biological Science, 2002, 384(1/2/3/4/5/6): 274-277. |
| [30] |
许大全, 张玉忠, 张荣铣. 植物光合作用的光抑制[J]. 植物生理学通讯, 1992(4): 237-243. XU Da-quan, ZHANG Yu-zhong, ZHANG Rong-xi. Photoinhibition of plant photosynthesis[J]. Plant Physiology Communications, 1992(4): 237-243. |
| [31] |
Cao C S, Xu J J, Zheng G Y, et al. Evidence for the role of transposons in the recruitment of cis-regulatory motifs during the evolution of C4 photosynthesis[J]. BMC Genomics, 2016, 17(1): 201. |