 2019, Vol. 38
2019, Vol. 38



2. 云南农业大学园林园艺学院, 昆明 650201
2. College of Horticulture and Landscape, Yunnan Agricultural University, Kunming 650201, China
农田土壤重金属污染已成为大家普遍关注的重大问题,耕作土壤中无论重金属含量高低,都会通过食物链逐级不可逆富集,严重影响人类健康发展[1-2]。因此,重金属污染土壤的治理已成为当前亟待解决的研究课题。由于目前土壤重金属污染源既有背景值,也有人为因素,因此不同重金属迁移转化和可给性不同,导致土壤中存在的重金属多呈现高度异质性斑块状分布[3-5]。植物对异质性营养斑块会产生不同的觅食行为,如杉木(Cunninghamia lanceolata)根系对不同磷斑块浓度与异质分布的阶段性产生不同的觅磷行为,土壤非均质性氮营养斑对玉米根系活力和吸收物质产生影响,营养斑中根密度一般是非营养斑处的10~15倍,沙生冰草(Agropyron desertorum)新生细根达2~5倍[6]。营养斑块对植物根系产生响应,胁迫斑块在超富集植物与作物间作体系中的效应如何,从重金属胁迫处理后两种植物体内含量差异显著可说明他们之间存在相关性[7-8],这种相关性是否与两种不同植物根系生长的趋向性有关有待继续探索。前人研究发现牛毛草对Hg吸收和异质性Pb胁迫对金丝草根长、体积和表面积产生显著影响[9]。本课题组前期对超富集植物小花南芥(Arabisalpina L. var. parviflora Franch)与玉米(Zea mays L.)、蚕豆(Vicia faba L.)间作修复Pb污染土壤做了大量研究,但对于重金属Pb胁迫下,间作体系如何影响小花南芥与玉米根系的形态学适应机制尚不清楚。
本团队前期研究发现,超富集植物小花南芥与玉米间作根系之间存在某种关联。土壤Pb含量为98.96 mg·kg-1的盆栽间作玉米与小花南芥发现,玉米Pb含量减少17.93%,小花南芥Pb含量增加9.62%[10];400 mg·kg-1 Pb胁迫水培处理后玉米体内Pb含量下降44%,小花南芥体内Pb含量增加75%,单作与间作根系形态差异较显著[11];而土壤Pb含量为600 mg·kg-1的大田间作蚕豆和小花南芥试验发现,与单作相比,间作体系下蚕豆Pb含量升高40.2%,小花南芥下降9.04%[12]。截至目前,本课题组研究还未能从本质意义上解释植物体内Pb含量的变化是否是根趋向行为引起。因此本研究设计垂直土壤Pb斑块处理为0、1000 mg·kg-1的根箱试验,以揭示超富集植物与作物间作体系下植物根系生长的趋向性形态学特征,为阐明间作体系下超富集植物小花南芥对Pb的富集和玉米对Pb的排斥提供科学依据。
1 材料与方法 1.1 试验材料供试小花南芥是本课题组2004年在云南省会泽县铅锌矿废弃地筛选的Pb超富集本土植物,其为十字花科南芥属植物[13],一年生草本,大多生长在废弃多年的矿渣上。试验采用野生种子育苗,挑选大小一致且籽粒饱满的小花南芥当年种子,种子用10%安替福民20倍液浸种20 min,蒸馏水清洗后,用镊子移入漂盘中,利用基质土进行漂浮育苗,每穴1~2粒繁育幼苗,繁育幼苗期间进行常规的浇水、光照等管理,培养的控制条件为:光(L):暗(D)=16 h:8 h,20±1 ℃,70% RH,出苗长至5片真叶后进行移栽。
供试玉米(Zea mays L.)品种为会单4号。挑选大小一致且籽粒饱满的种子,用10%双氧水表面消毒10 min,蒸馏水清洗后,直接播种于根箱内。
1.2 试验方法土壤Pb胁迫处理:用分析纯试剂Pb(NO3)2配制(以纯Pb2+计1000 mg·kg-1)成500 mL溶液,然后一次性均匀浇灌在土壤中,对照无Pb处理用同样的蒸馏水均匀浇灌在土壤中,1000 mg·kg-1 Pb(NO3)2处理所有土壤,搅拌平衡1个月待用。本试验设计的Pb胁迫浓度参照陈建军等[10]的文献,保证Pb胁迫不影响玉米和小花南芥的正常生长发育。
2015年2月23日在云南农业大学化学楼旁温室大棚进行根箱试验,根箱为透明有机玻璃制作,高60 cm×宽40 cm×厚2 cm。土壤为同质土壤Pb处理0、1000 mg·kg-1及异质性土壤Pb处理斑块0、1000 mg· kg−1,异质性土壤处理时在中间用塑料片(1 mm)隔开,两边分别装入0 mg·kg-1和1000 mg·kg-1Pb处理,根箱两边分别种植一株植物,移栽时注意保证根系自然伸展,且正好处于一侧的中心位置。种植完成后抽出塑料片,为了防止异质土壤中Pb2+迁移到另外一侧,利用250 mL注射器从两侧缓慢注射水,保证水恰好浸透到根箱中间位置。试验设3种种植模式,分别为玉米单作、小花南芥单作、玉米与小花南芥间作。每个处理3次重复,合计30盆。根箱试验具体设计见图 1。

|
图 1 土培试验装置 Figure 1 Diagram for experimental unit |
2015年5月23日从根箱固定架上取下根箱,水平放置于地面上,取下固定夹子,有机玻璃板移走一面,从两株植物中间向两边依次0~10、10~20 cm刀片分割,取出每个区域内的植物根系并做好标记后备用。
1.4 根系形态测定将按不同距离采集的根系鲜样,用自来水冲洗数次,再用蒸馏水冲洗1次,然后用EPSON PERFECTIONV700扫描仪扫描,用根系分析软件WinRHIZO-Pro 2013(Regent Instruments Inc)对扫描图片进行分析。依据Costa等[14]的方法统计玉米和小花南芥的根表面积、径级根系指标。用电子天平称量根系鲜质量。
1.5 数据处理与统计分析 1.5.1 竞争关系分析以根系生物量为基础,采用相对关系指数RII(Relative interaction index)[15-16]衡量间作条件下作物的竞争能力:
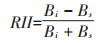
|
式中:Bi表示小花南芥与玉米间作根与根之间作用条件下的生物量;Bs表示小花南芥或玉米在单作种植条件下的生物量。当RII>0时,表示玉米(小花南芥)在竞争中得到的互利效应大于受到的竞争效应,当RII < 0时,则竞争效应大于互利效应,当RII=0时,玉米(小花南芥)在间作中没有受到影响。RII值越大,表明相对竞争能力越强。
1.5.2 小花南芥趋Pb和玉米避Pb超富集植物和作物间作在同一空间土壤中,各自为适应土壤中不同元素的异质性,其根系诱导出一系列的生理和表型可塑性。Pb超富集植物小花南芥以趋Pb为主,而Pb为玉米的胁迫物质,玉米主要以避Pb为主。因此,其中增加Pb处理斑块内的根系生长被认为是小花南芥对异质性Pb的适应性响应,采用Pb处理斑块内与无Pb处理斑块内根系生物量之比来衡量趋Pb相对指数。比值越高,其趋Pb相对指数越高;对于玉米,比值越低,其避Pb相对指数越高。
1.5.3 数据分析数据处理使用Excel Microsoft,利用DPS数据处理系统中Duncan新复极差法进行差异显著性统计分析。分析竞争关系时以作物和Pb异质性为固定因素,分析植物系趋向性时以作物和种植模式为固定因素。
2 结果与分析 2.1 间作体系对Pb斑块处理植物根系生物量的影响 2.1.1 玉米根系生物量同质土壤Pb处理为0 mg·kg-1和1000 mg·kg-1时,同侧间作玉米根系在0~10 cm处生物量比单作分别增加48.83%和24.73%,0~10 cm与10~20 cm相比,间作玉米根系生物量分别增加43.12%和31.73%,差异较显著,同侧在10~20 cm处间作玉米与单作生物量差异不显著。单作玉米根系生物量A侧和B侧在0~ 10 cm和10~20 cm差异均不显著。从表 1中A侧和B侧根系总生物量看,Pb胁迫处理后单作和间作生物量均下降,说明受Pb污染的土壤会直接影响植物的生长发育。对同质土壤0~10 cm处两因素比较分析可知,种植模式和Pb处理交叉因素对玉米根系生物量产生极显著影响(F=24.89,P=0.001)。
|
|
表 1 间作体系对异质土壤Pb处理玉米根系生物量的影响 (mg·kg-1) Table 1 Effects of Pb heterogeneous stresses on roots biomass of intercropped maize in the intercropping system (mg·kg-1) |
异质土壤Pb处理斑块为0 mg·kg-1和1000 mg· kg−1时,玉米种植在B侧Pb胁迫处理斑块为1000 mg· kg−1时,间作玉米0~10 cm处根系比单作显著增加28.40%,10~20 cm处差异不显著,间作体系下,0~10 cm处比10~20 cm处玉米生物量显著增加37.76%;玉米种植在B侧Pb胁迫处理斑块为0 mg·kg-1时,0~10 cm处间作玉米根系比单作显著减少11.77%,10~20 cm处显著增加27.01%,单作玉米生物量在0~10 cm处比10~20 cm处显著增加7.94%,间作玉米生物量在0~10 cm处比10~20 cm处显著减少25.03%,说明异质性土壤间作改变玉米根系的生物量集中在玉米与小花南芥接触的微区域内。对异质土壤0~10 cm处两因素比较分析可知,Pb处理因素对玉米根系生物量产生极显著影响(F=22.75,P=0.001)
2.1.2 小花南芥根系生物量同质土壤Pb处理为0 mg·kg-1时,间作小花南芥与单作在同侧0~10 cm和10~20 cm处根系生物量差异不显著,但间作体系下小花南芥总生物量平均值比单作增加0.5倍;同质土壤Pb处理为1000 mg·kg-1时,间作小花南芥与单作在同侧0~10 cm和10~20 cm处根系生物量显著增加90.12%和100.72%,间作体系下0~10 cm处小花南芥总生物量平均值比单作增加0.95倍。对同质土壤0~10 cm处两因素比较分析可知,种植模式因素对小花南芥根系生物量产生极显著影响(F=81.68,P < 0.001)。
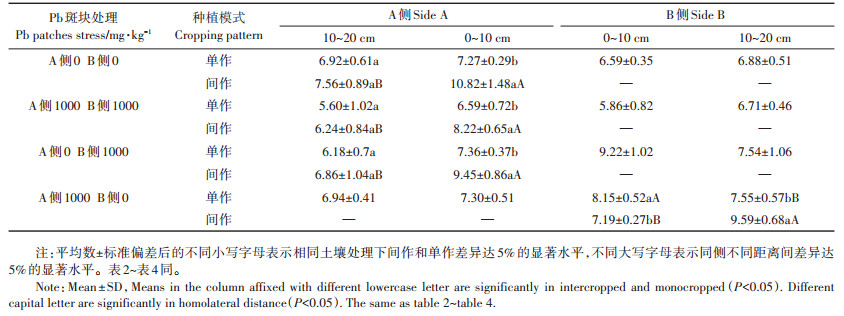
异质土壤Pb处理斑块为0 mg·kg-1和1000 mg· kg−1时,小花南芥种植在B侧Pb胁迫处理斑块为1000 mg·kg-1时,0~10 cm和10~20 cm处间作小花南芥根系比单作显著增加55.39%和48.60%,间作体系下小花南芥总生物量平均值比单作增加47%。小花南芥种植在B侧Pb胁迫处理斑块为0 mg·kg-1时,0~ 10 cm和10~20 cm处间作小花南芥根系比单作显著增加68.89%和83.33%。从表 2看出,超富集植物小花南芥根系生物量在Pb胁迫和间作处理后与无Pb胁迫和单作处理差异显著。对异质土壤0~10 cm处两因素比较分析可知,种植模式因素对小花南芥根系生物量产生极显著影响(F=60.15,P < 0.001)。
|
|
表 2 间作体系对异质土壤Pb处理小花南芥根系生物量的影响 (mg·kg-1) Table 2 Effects of Pb heterogeneous stresses on roots biomass of Arbais alpine in the intercropping system (mg·kg-1) |
如图 2所示,玉米种植于同质土壤0 mg·kg-1一侧时,Pb异质性显著降低玉米在0~10 cm处两作物间的竞争,而10~20 cm处同质和异质土壤玉米在竞争中得到的互利效应大于受到的竞争效应,且异质土壤上玉米根系的互利效应与同质土壤相比显著增加4.25倍。玉米种植于同质土壤1000 mg·kg-1一侧时,玉米根系均表现为互利效应,但同质与异质土壤在同距离处理之间变化不显著。

|
图中不同小写字母表示同一Pb浓度不同处理间差异达5%的显著水平。图 3同 Means in the figure affixed with different lowercase letter are significantly between different planting modes under the same Pb treatment (P < 0.05). The same as figure 3 图 2 Pb异质性对间作玉米根系竞争关系的影响 Figure 2 Effects of Pb heterogeneous stresses on roots competitive relation of intercropped maize |

|
图 3 Pb异质性对间作小花南芥根系竞争关系的影响 Figure 3 Effects of Pb heterogeneous stresses on roots competitive relation of intercropped Arbais alpine |
如图 3所示,Pb处理为0 mg·kg-1时,小花南芥在同质土壤0~10 cm处和异质土壤0~10 cm和10~20 cm处相对关系指数平均为0.40,同质土壤10~20 cm处时相对关系指数为1.19,极显著增加了1.98倍。Pb处理为1000 mg·kg-1时,小花南芥相对关系指数平均为0.27,处理间差异不显著。从图 2和图 3可以看出,Pb处理为0 mg·kg-1时小花南芥根与玉米根间互利效应大于1000 mg·kg-1Pb胁迫处理。
2.3 植物根系对Pb斑块趋向性分析 2.3.1 玉米根系避Pb相对指数分析如图 4所示,单作玉米根系在同质土壤中的平均避Pb相对指数为0.86,异质土壤在10~20 cm处避Pb相对指数为0.51,相比同质土壤显著下降41.17%;间作玉米根系在同质土壤中的平均避Pb相对指数为0.77,在异质土壤0~10 cm处避Pb相对指数为1.31,相比同质土壤显著增加70.86%。同质土壤中玉米根系在10~20 cm处避Pb相对指数显著降低,异质性土壤中玉米根系在0~10 cm处避Pb相对指数显著增高,说明Pb胁迫斑块改变了玉米根系的避Pb相对指数。

|
图中不同小写字母表示同一种植方式不同处理间差异达5%的显著水平。图 5同 Means in the figure affixed with different lowercase letter are significantly in the same model (P < 0.05). The same as figure 5 图 4 根间作用对玉米根系避Pb相对指数的影响 Figure 4 Effects of root interaction on roots prevent Pb precision of intercropped maize |

|
图 5 根间作用对小花南芥根系趋Pb相对指数的影响 Figure 5 Effects of root interaction on roots foraging Pb precision of intercropped Arbais alpine |
如图 5所示,单作小花南芥在同质土壤中的平均趋Pb相对指数为5.81,在异质土壤10~20 cm处趋Pb相对指数为8.51,相比同质土壤显著增加46.47%;间作小花南芥在同质土壤0~10 cm处的趋Pb相对指数为2.48,在异质土壤中平均趋Pb相对指数为7.89,相比同质土壤0~10 cm处显著增加2.18倍。单作小花南芥在异质土壤10~20 cm处趋Pb相对指数显著增高,间作小花南芥除了在同质土壤0~10 cm处趋Pb相对指数显著下降外,其他趋Pb相对指数差异不显著,说明小花南芥根系具有寻找Pb的能力。
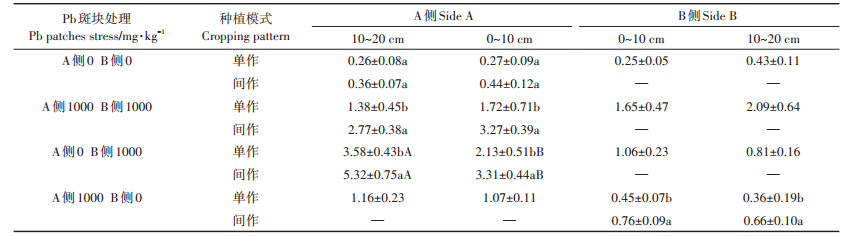
2.4 间作体系对Pb斑块处理植物根表面积的影响 2.4.1 对玉米根表面积的影响如表 3所示,同质土壤Pb处理为0 mg·kg-1时,间作玉米根表面积在0~10 cm处与单作相比显著增加42.89%,单作玉米0~10 cm较10~20 cm处显著降低30.19%;同侧间作玉米根表面积在0~10 cm处与10~ 20 cm处差异不显著。Pb胁迫处理为1000 mg·kg-1时,间作玉米在0~10 cm处与单作相比显著增加52.63%,且0~10 cm处比10~20 cm处显著增加0.75倍,间作对根系表面积影响主要集中在0~10 cm处微区域内。
|
|
表 3 间作体系对异质土壤Pb处理玉米根表面积的影响 Table 3 Effects of Pb heterogeneous stresses on root surface area of intercropped maize in the intercropping system |
异质土壤Pb处理斑块为0 mg·kg-1和1000 mg· kg−1时,玉米种植在B侧Pb胁迫处理斑块为1000 mg· kg−1时,0~10 cm处间作玉米根表面积比单作显著增加32.90%,A侧间作玉米根表面积在0~10 cm处比10~20 cm处显著增加30.08%。玉米种植在B侧Pb胁迫处理斑块为0 mg·kg-1时,0~10 cm和10~20 cm处间作玉米根表面积与单作相比显著增加35.01%和36.02%。异质土壤Pb斑块处理后,间作体系对玉米根表面积影响较显著。对同质(F=15.70,P=0.004)和异质(F=22.41,P=0.001)土壤0~10 cm处两因素比较分析可知,种植模式和Pb处理因素对玉米根表面积产生极显著影响。
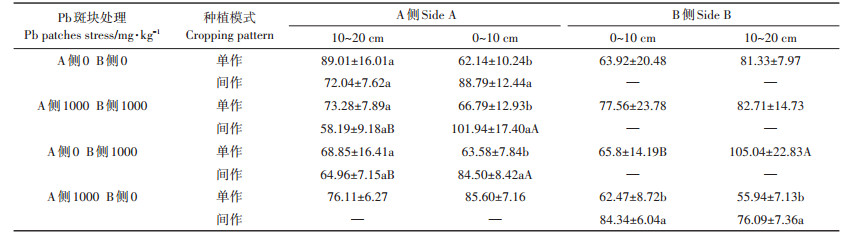
2.4.2 对小花南芥根表面积的影响如表 4所示,同质土壤Pb处理为0 mg·kg-1时,间作小花南芥根表面积在0~10 cm处与单作相比显著增加45.35%;同侧间作小花南芥根表面积在0~10 cm处比10~20 cm处显著增加81.02%。Pb胁迫处理为1000 mg·kg-1时,间作小花南芥在0~10 cm和10~20 cm处与单作相比显著增加31.53%和40.09%。A侧和B侧的单作小花南芥0~10 cm和10~20 cm处根表面积差异均不显著。对同质土壤0~10 cm处两因素比较分析可知,种植模式和Pb处理因素对小花南芥根表面积产生极显著影响(F=17.07,P=0.003)。
|
|
表 4 间作体系对异质土壤Pb处理小花南芥根表面积的影响 Table 4 Effects of Pb heterogeneous stresses on root surface area of intercropped Arbais alpine in the intercropping system |
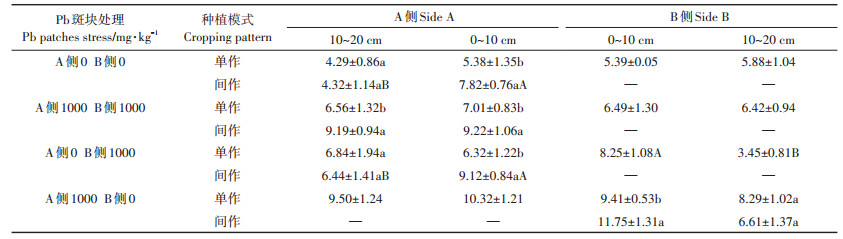
异质土壤Pb处理斑块为0 mg·kg-1和1000 mg· kg−1时,小花南芥种植在B侧Pb胁迫处理斑块为1000 mg·kg-1时,0~10 cm处间作小花南芥根表面积比单作显著增加44.30%,A侧间作小花南芥根表面积在0~10 cm处较10~20 cm处显著增加41.61%。小花南芥种植在B侧Pb胁迫处理斑块为0 mg·kg-1时,0~10 cm间作小花南芥根表面积与单作相比显著增加24.87%,且0~10 cm处较10~20 cm处显著增加77.76%。异质土壤Pb斑块处理后,间作体系下小花南芥根表面积发生显著变化区域主要在0~10 cm交界处。对异质(F=12.58,P=0.008)土壤0~10 cm处两因素比较分析可知,种植模式因素对小花南芥根表面积产生极显著影响。
2.5 玉米和小花南芥间作体系对Pb斑块处理植物根表面积径级的影响根系表面积的大小影响根系吸收离子的强度。从表 5可知,玉米间作和单作根系表面积在d4径级区间占总根系表面积的比例最大,分别为27.01%和40.73%,并且间作下表面积比单作显著下降了36.98%(P < 0.05);d1和d3径级玉米间作表面积比单作显著增加了26%和32%(P < 0.05);间作体系下玉米根系在各径级间表面积分布范围为11.95~25.88 cm2,平均为18.05 cm2;单作分布范围为9.41~41.07 cm2,平均为20.94 cm2,说明间作体系下玉米根系表面积分布较均匀,这有利于玉米根系从土壤中均衡吸收离子。
|
|
表 5 间作体系对小花南芥和玉米根表面积径级的影响 (cm2) Table 5 Effects of root surface diameter classes on intercropped Arbais alpine and maize (cm2) |
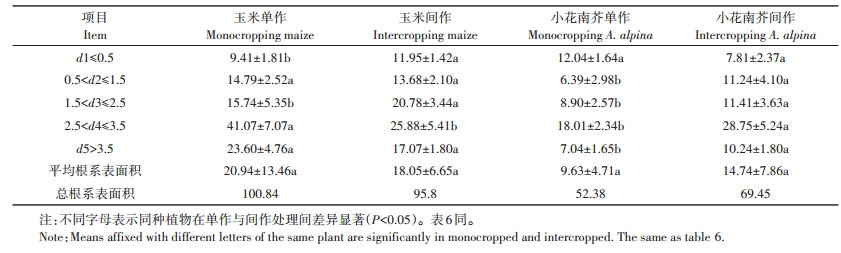
小花南芥单作在d1、d2、d3、d4、d5 5个径级区间占总表面积的比例分别为22.98%、12.19%、16.99%、34.38%、13.44%,间作分别为11.25%、16.18%、16.43%、41.39%、14.74%,除了d1径级根系表面积间作与单作差异不显著外,其余径级间作与对照差异均显著,间作体系下分别增加了76%、28%、60%、45%,说明间作体系促进小花南芥根系的生长发育。
从表 5知,间作体系下根系表面积比单作显著增加,主要影响玉米和小花南芥根系生长状态的径级在1.5 < d3≤2.5和2.5 < d4≤3.5区间。
2.6 玉米和小花南芥间作体系对Pb富集特征的影响间作体系根-根之间的相互作用,不仅会改变植物对营养物质的竞争,同时也影响非营养元素的吸收累积。从表 6可知,玉米间作与单作相比,地上部和地下部Pb的累积量分别显著降低了38.98%和17.80%(P < 0.05),并且地上部的降幅大于地下部;玉米间作转运系数为0.32,单作为0.43,间作体系使转运系数较单作下降了25.58%。
|
|
表 6 间作体系下玉米和小花南芥富集特征 Table 6 Effects of the accumulation of lead in the upper and underground parts on intercropped Arbais alpine and maize |

小花南芥间作与单作相比地上部显著增加了45.64%,相反地下部下降了18.99%;小花南芥间作转运系数为2.25,单作为1.25,间作体系使转运系数较单作增加了80%。
从以上数据可知,间作体系下玉米转运到地上部的Pb量减少,而小花南芥地上部的Pb量却增加,说明玉米与小花南芥间作显著改变了Pb在两种植物地下部和地上部的累积量。
3 讨论 3.1 小花南芥趋Pb和玉米避Pb相对指数分析间作条件下,植物根系趋向性的改变通常是植物适应外界异质土壤环境的策略。土壤营养斑块中植物根趋向营养一边,有助于促进植物对营养的吸收,增强竞争优势[17-19],但在土壤氮斑块处理玉米和马铃薯时,玉米生物量和竞争能力都增加,马铃薯并未获利[15]。说明植物根系趋向性不一定能增强竞争优势[20-22],趋向行为本身是一个消耗能量的过程,在竞争过程中将有限的能量用于根系的趋向性生长行为可能降低净生物量的增加[23]。在本研究中,单作玉米异质土壤趋向性指数比同质土壤低,而间作玉米却相反;小花南芥单作和间作异质土壤斑块趋向指数均比同质土壤高;玉米和小花南芥生物量在不同处理下均显著增加。表明吸收不同营养物质的植物间作可以通过根间相互作用在异质性Pb土壤环境下提高植物的生物量,这与已有的异质性氮环境可以提高玉米和马铃薯总生产力的结论存在一致性[15]。异质性的营养斑块促进植物多样性与生产力的正相关性[24],这可能与植物根系的觅养能力紧密相关[25],超富集植物小花南芥以Pb为主要的营养物质,异质性Pb斑块中小花南芥根系采取主动吸收途径获取营养成分,而作物玉米在异质性Pb斑块中会避开Pb斑块获取自身需要的营养物质。超富集植物与作物间作下植物不同根系在异质性斑块上有可能通过根系表型的可塑性改变根趋向行为[9]。
3.2 小花南芥与玉米根系竞争关系分析已有研究证明间作植物根系在斑块土壤处理下通过表型可塑性来选择更加适合的区域生产而增加资源的获取量。苦豆子根系在无竞争时,采用“广布策略”,在有竞争者时,根系受到抑制而分布在较小范围内,且土壤均质时,未重叠区域的根系比例较高;资源斑块出现时,斑块中的重叠部分根系比例较高,由于资源丰富而不受重叠的影响;斑块在边缘时,原位植物更容易获得丰富的资源,因此趋向中心一侧的根系[26]。本研究小花南芥与玉米间作时,异质性Pb处理下间作玉米在0~10 cm处表现出显著的竞争效应,同质和异质性Pb处理小花南芥在10~20 cm处表现为互利效应,这与苦豆子根系特征趋势一致。两因素分析显示,交叉因素对玉米根生物量影响极显著,种植模式对小花南芥根生物量影响极显著。Cahill等[27]研究苘麻(A. theophrasti)根系觅养能力时发现,没有竞争时根系水平生长幅度不受营养斑块的影响,有竞争时则在土壤同质性营养环境中受到显著抑制,而在异质性营养斑块环境中影响不显著。在本研究中,同质性和异质性Pb处理斑块间作小花南芥和玉米生物量都表现出正效应。Cahill等[27]从种群水平提出植物对异质性元素和间作竞争作出响应的同时,也对邻近的竞争者作出非资源性介导的响应。吴开贤等[15]从群落水平验证植物种间根-根作用,发现其可提高玉米和马铃薯对异质性氮的觅养精确度,激发其觅养能力,但该研究还发现异质性氮并未增加根间竞争强度。也有研究表明,间作植物根-根作用导致土壤有效养分降低,有限空间内两种植物为生存可能提高觅养能力[28-29]。本研究中异质性玉米Pb处理增加了玉米根系的竞争强度,而小花南芥在竞争中获利。说明植物根系能够整合异质性养分和邻近者的根间作用信息,植物将根系分泌物作为信号物质来区分邻近植株的遗传身份以作出差异响应[30]。当植物根系界面面对遗传信息相近或相同植株时,植株觅养精确度可能降低,当面对亲缘关系较远的植物时,觅养精确度则可能提高,植物对养分的吸收遵循Hamilton的亲缘选择理论(Kin selection theory)。超富集植物小花南芥与低富集作物玉米属于亲缘关系相差甚远的两个物种,这可能是解释富集Pb差别的原因之一。小花南芥对Pb的需求量较大,因此吸收速率较快,斑块内的养分发出下降趋势的信号后,玉米感知到这一信号,玉米的觅养可能转向没有竞争对象的低养分斑块。从而玉米根系表型形态发生变化,根系觅养行为将选择未被“占领”区域[31]。这有待于进一步从原位研究超富集植物与低富集作物感知养分异质性环境和资源的瞬时性动态行为[32],尤其是对根系分泌物发出觅食信号做出差异响应,这可揭示小花南芥与玉米对Pb的趋向性行为。
3.3 根系形态变化的适应对策植物根系形态对外界环境的变化较为敏感,通常根据表型可塑性变化判断植物根系对异质性环境的适应对策,特别是在同一空间生长的不同物种之间根系形态适应对策差异较大时。有研究表明植株通过根的形态学和生理学的改变来适应不利环境的胁迫[33-34]。通过通径分析根系形态指标与氮吸收效率,发现根长和根体积对氮吸收效率有直接影响作用,其他指标通过根长和根体积产生间接作用[35]。金丝草在土壤Pb斑块中通过根长、根体积和表面积增大来增加与土壤Pb的接触,从而提高对Pb的富集能力[9]。磷饥饿环境下磷高效基因型杉木的根表面积和根体积显著大于正常供磷的斑块,植物根表面积和体积增大,缩短了磷离子扩散到植物根的距离[36]。本研究中,同质性和异质性Pb处理土壤,间作玉米和小花南芥表面积在0~10 cm处显著增加,间作模式和Pb处理两因素分析表明,间作模式和Pb处理极显著影响玉米根表面积,而间作模式极显著影响小花南芥根表面积。这说明资源异质性的改变可能是一种综合变化,验证了相邻的植物之间存在非资源性介导的响应。上述数据说明表型可塑性能反映间作植物根系与其生境资源条件相适应的密切关系。进一步研究表明,根系形态的变化与植物体内特定基因的表达调控和内源激素存在相关性[37],揭示玉米和小花南芥间作表型改变可能需要从两者基因表达差异方面深入探索。
3.4 小花南芥和玉米对Pb富集特征的影响Pb是植物生长的非必需元素,其对植物产生胁迫时,会使植物的形态学特征发生变化,甚至导致植物因不适应胁迫而死亡。但有研究发现,Pb超富集植物在受到胁迫时会产生适应机制,如金丝草根系遇到Pb胁迫时,会选择向两侧生长,当两侧根系生长遇阻时,也具有适应Pb胁迫斑块生长的能力[9],这说明不同植物对Pb胁迫的适应机制存在差异。本文中小花南芥与玉米间作在Pb斑块胁迫下,两种植物根系趋向行为发生显著差异,推测可能存在两种机制:一是小花南芥和玉米对Pb排斥机制差异,间作根系分泌柠檬酸促使小花南芥根系吸收富集Pb[11],小花南芥富集的Pb可能通过细胞壁和细胞外的碳水化合物把Pb排斥在细胞质外,而草酸与Pb2+结合阻止玉米根系吸收[11, 38];二是防御机制差异,小花南芥过氧化酶活性增强,导致根趋向有Pb斑块生长,玉米超氧化物歧化酶活性增强,导致根趋向无Pb斑块生长[39],植物根的趋向行为引起Pb富集特征的变化。
4 结论(1)Pb斑块处理根箱条件下,间作小花南芥在0~ 10 cm处根系趋向有Pb斑块生长,生物量和表面积显著增加;间作玉米根系趋向无Pb斑块生长,根相对关系指数显著下降。
(2)间作降低玉米转运系数,增加小花南芥转运系数;根表面积径级在1.5 < d3≤2.5时显著影响间作小花南芥和玉米对Pb胁迫的响应。
| [1] |
Ameh E G, Akpah F A. Heavy metal pollution indexing and multivariate statistical evaluation of hydrogeochemistry of River PovPov in Itakpe Iron-Ore mining area, Kogi State, Nigeria[J]. Advances in Applied Science Research, 2011, 2(1): 33-46. |
| [2] |
李瑞琴, 于安芬, 白滨, 等. 甘肃中部高原露地菜田土壤重金属污染及潜在生态风险分析[J]. 农业环境科学学报, 2013, 32(1): 103-110. LI Rui-qin, YU An-fen, BAI Bin, et al. Analysis on current situation and potential ecological risk and the characteristic of heavy metals pollution of soil in the central plateau of Gansu Province, China[J]. Journal of Agro-Environment Science, 2013, 32(1): 103-110. |
| [3] |
丛源, 陈岳龙, 杨忠芳, 等. 北京市农田土壤重金属的化学形态及其对生态系统的潜在危害[J]. 土壤, 2009, 41(1): 39-43. CONG Yuan, CHEN Yue-long, YANG Zhong-fang, et al. Chemical forms of heavy metals in soils and potential hazards to ecosystem in Beijing farmlands[J]. Soils, 2009, 41(1): 39-43. |
| [4] |
王亚宇, 钱翌, 朱建雯, 等. 乌鲁木齐市表层土壤Pb的化学形态分布及潜在生态危害评价[J]. 环境科学研究, 2008, 21(5): 106-111. WANG Ya-yu, QIAN Yi, ZHU Jian-wen, et al. Chemical speciation of Pb and its potential ecological risk in urban superficial soils of Urumqi City[J]. Research of Environmental Sciences, 2008, 21(5): 106-111. |
| [5] |
Banata K M, Howarib F M, AIhamard A A. Heavy metals in urban soils of central Jordan:Should we worry about their environmental risks?[J]. Environmental Research, 2005, 97(3): 258-273. DOI:10.1016/j.envres.2004.07.002 |
| [6] |
邹显花, 吴鹏飞, 贾亚运, 等. 杉木根系对不同磷斑块浓度与异质分布的阶段性响应[J]. 植物营养与肥料学报, 2016, 22(4): 1056-1063. ZOU Xian-hua, WU Peng-fei, JIA Ya-yun, et al. Periodical response of Chinese fir root to the phosphorus concentrations in patches and heterogeneous distribution in different growing stages[J]. Journal of Plant Nutrition and Fertilizer, 2016, 22(4): 1056-1063. |
| [7] |
秦丽, 祖艳群, 湛方栋, 等. 续断菊与玉米间作对作物吸收积累镉的影响[J]. 农业环境科学学报, 2013, 32(3): 471-477. QIN Li, ZU Yan-qun, ZHAN Fang-dong, et al. Absorption and accumulation of Cd by Sonchus asper L. Hill. and maize in intercropping systems[J]. Journal of Agro-Environment Science, 2013, 32(3): 471-477. |
| [8] |
湛方栋, 陈建军, 秦丽, 等. 镉铅污染的玉米秸秆还田对蚕豆生长、养分和镉铅含量的影响[J]. 农业环境科学学报, 2016, 35(4): 661-668. ZHAN Fang-dong, CHEN Jian-jun, QIN Li, et al. Effects of applying Cd/Pb contaminated maize stalks on growth and nutrient and Cd and Pb content of fababean[J]. Journal of Agro-Environment Science, 2016, 35(4): 661-668. |
| [9] |
侯晓龙, 刘爱琴, 蔡丽平, 等. 异质Pb胁迫对超富集植物金丝草根系生长及生物量分配的影响[J]. 农业环境科学学报, 2013, 32(11): 2145-2150. HOU Xiao-long, LIU Ai-qin, CAI Li-ping, et al. Effects of heterogeneous Pb stress on root growth and biomass allocation of Pogonatherum crinitum[J]. Journal of Agro-Environment Science, 2013, 32(11): 2145-2150. DOI:10.11654/jaes.2013.11.007 |
| [10] |
陈建军, 修珩, 李元. 小花南芥与玉米间作对Pb的吸收累积特征[J]. 环境科学与技术, 2016, 39(11): 63-67. CHEN Jian-jun, XIU Heng, LI Yuan. Absorption and accumulation of Pb with Arabis alpina L. and Zea mays in intercropping system[J]. Environmental Science & Technology, 2016, 39(11): 63-67. |
| [11] |
王吉秀, 湛方栋, 李元, 等. 铅胁迫下小花南芥与玉米间作对根系分泌物有机酸的影响[J]. 中国生态农业学报, 2016, 24(3): 365-372. WANG Ji-xiu, ZHAN Fang-dong, LI Yuan, et al. Effects of Arabisalpina L. var. parviflora Franch and Zea mays L. intercropping system on root-exudated organic acids under lead stress[J]. Chinese Journal of Eco-Agriculture, 2016, 24(3): 365-372. |
| [12] |
陈兴, 郭先华, 祖艳群, 等. 蚕豆与小花南芥间作体系中Cd, Pb在植物中的累积特征[J]. 云南农业大学学报(自然科学), 2016, 31(1): 167-172. CHEN Xing, GUO Xian-hua, ZU Yan-qun, et al. Accumulation characteristics of Cd and Pb in broad bean/Arabis alpina intercropping system[J]. Journal of Yunnan Agricultural University(Natural Science), 2016, 31(1): 167-172. |
| [13] |
Zu Y Q, Li Y, Christuan S, et al. Accumulation of Pb, Cd, Cu and Zn in plants and hyperaccumulator choice in Lanping lead-zinc mine area, China[J]. Environment International, 2004, 30(4): 567-576. DOI:10.1016/j.envint.2003.10.012 |
| [14] |
Costa C, Dwyer L M, Dwyer M, et al. A sampling method for measurement of large root systems with scanner-based image analysis[J]. Agron J, 2000, 92(4): 621-627. DOI:10.2134/agronj2000.924621x |
| [15] |
吴开贤, 安曈昕, 范志伟, 等. 根间相互作用对玉米与马铃薯响应异质氮的调控[J]. 生态学报, 2015, 35(2): 508-516. WU Kai-xian, AN Tong-xin, FAN Zhi-wei, et al. Effects of root interactions on the response of maize and potato to heterogeneous nitrogen[J]. Acta Ecologica Sinica, 2015, 35(2): 508-516. |
| [16] |
Armas C, Ordiales R, Pugnaire F I. Measuring plant interactions:A new comparative index[J]. Ecology, 2004, 85(10): 2682-2686. DOI:10.1890/03-0650 |
| [17] |
Robinson D, Hodge A, Griffiths B S, et al. Plant root proliferation in nitrogen-rich patches confers competitive advantage[J]. Proceedings of the Royal Society of London, Series B:Biological Sciences, 1999, 266(1418): 431-437. DOI:10.1098/rspb.1999.0656 |
| [18] |
Hodge A. The plastic plant:Root responses to heterogeneous supplies of nutrients[J]. New Phytologist, 2004, 162(1): 9-24. DOI:10.1111/j.1469-8137.2004.01015.x |
| [19] |
Nakamura R, Kachi N, Suzuki J I. Root growth and plant biomass in Lolium perenne exploring a nutrient-rich patch in soil[J]. Journal of Plant Research, 2008, 121(6): 547-557. DOI:10.1007/s10265-008-0183-7 |
| [20] |
Jackson R B, Manwaring J H, Caldwell M M. Rapid physiological adjustment of roots to localized soil enrichment[J]. Nature, 1990, 344(6261): 58-60. DOI:10.1038/344058a0 |
| [21] |
Maestre F T, Reynolds J F. Amount or pattern? Grassland responses to the heterogeneity and availability of two key resources[J]. Ecology, 2007, 88(2): 501-511. DOI:10.1890/06-0421 |
| [22] |
Jansen C, Van Kempen M M L, Bögemann G M, et al. Limited costs of wrong root placement in Rumex palustris in heterogeneous soils[J]. New Phytologist, 2006, 171(1): 117-126. DOI:10.1111/j.1469-8137.2006.01733.x |
| [23] |
Forde B, Lorenzo H. The nutritional control of root development[J]. Plant and Soil, 2001, 232(1): 51-68. |
| [24] |
Tylianakis J M, Rand T A, Kahmen A, et al. Resource heterogeneity moderates the biodiversity-function relationship in real world ecosystems[J]. PLoS Biology, 2008, 6(5): e122. DOI:10.1371/journal.pbio.0060122 |
| [25] |
García Palacios P, Maestre F T, Gallardo A. Soil nutrient heterogeneity modulates ecosystem responses to changes in the identity and richness of plant functional groups[J]. Journal of Ecology, 2011, 99(2): 551-562. |
| [26] |
郝鹏, 李景文, 丛日春, 等. 苦豆子根系对土壤异质性和竞争者的响应[J]. 北京林业大学学报, 2012, 34(5): 94-99. HAO Peng, LI Jing-wen, CONG Ri-chun, et al. Response of arid plant Sophora alopecuroides to soil heterogeneity and competitors[J]. Journal of Beijing Forestry University, 2012, 34(5): 94-99. |
| [27] |
Cahill J F, Mc Nickle G G, Haag J J, et al. Plants integrate information about nutrients and neighbors[J]. Science, 2010, 328(5986): 1657. DOI:10.1126/science.1189736 |
| [28] |
Bilbrough C J, Caldwell M M. The effects of shading and N status on root proliferation in nutrient patches by the perennial grass Agropyron desertorum in the field[J]. Oecologia, 1995, 103(1): 10-16. DOI:10.1007/BF00328419 |
| [29] |
Maestre F T, Reynolds J F. Small-scale spatial heterogeneity in the vertical distribution of soil nutrients has limited effects on the growth and development of Prosopis glandulosa seedlings[J]. Plant Ecology, 2006, 183(1): 65-75. DOI:10.1007/s11258-005-9007-1 |
| [30] |
Fang S, Gao X, Deng Y, et al. Crop root behavior coordinates phosphorus status and neighbors:From field studies to three-dimensional in situ reconstruction of root system architecture[J]. Plant Physiology, 2011, 155(3): 1277-1285. DOI:10.1104/pp.110.167304 |
| [31] |
Semchenko M, Zobel K, Hutchings M J. To compete or not to compete:An experimental study of interactions between plant species with contrasting root behaviour[J]. Evolutionary Ecology, 2010, 24(6): 1433-1445. DOI:10.1007/s10682-010-9401-6 |
| [32] |
Shemesh H, Ovadia O, Novoplansky A. Prioritized contingencies:Context-dependent regeneratory effects of grazer saliva[J]. Plant Ecology, 2012, 213(1): 167-174. DOI:10.1007/s11258-011-9968-1 |
| [33] |
于兆国, 张淑香. 不同磷效率玉米自交系根系形态与根际特征的差异[J]. 植物营养与肥料学报, 2008, 14(6): 1227-1231. YU Zhao-guo, ZHANG Shu-xiang. Root configuration and rhizosphere characteristics of different maize inbred lines with contrasting P efficiency[J]. Journal of Plant Nutrition and Fertilizer Science, 2008, 14(6): 1227-1231. DOI:10.3321/j.issn:1008-505X.2008.06.031 |
| [34] |
刘梅, 吴广俊, 路笃旭, 等. 不同年代玉米品种氮素利用效率与其根系特征的关系[J]. 植物营养与肥料学报, 2017, 23(1): 71-82. LIU Mei, WU Guang-jun, LU Du-xu, et al. Improvement of nitrogen use efficiency and the relationship with root system characters of maize cultivars in different years[J]. Journal of Plant Nutrition and Fertilizer Science, 2017, 23(1): 71-82. |
| [35] |
屈佳伟, 高聚林, 王志刚, 等. 玉米根系特征的基因型差异及与氮吸收效率的关系[J]. 玉米科学, 2016, 24(2): 72-78. QU Jia-wei, GAO Ju-lin, WANG Zhi-gang, et al. Genotype differences in root characteristics by maize and its relations to nitrogen uptake efficiency[J]. Journal of Maize Sciences, 2016, 24(2): 72-78. |
| [36] |
Ma Q F, Ren G Z. Phosphorus acquisition and wheat growth are influenced by shoot phosphorus status and soil phosphorus distribution in a split-root system[J]. Journal of Plant Nutrition and Soil Science, 2008, 171(2): 266-271. DOI:10.1002/jpln.200700183 |
| [37] |
李丽杰, 顾万荣, 王泳超, 等. DCPTA浸种对寒地玉米种子萌发及根系生长的影响[J]. 作物杂志, 2015(4): 152-156. LI Li-jie, GU Wan-rong, WANG Yong-chao, et al. Effects of seed soaking with DCPTA solution on seed germination and root formation of spring maize in cold region[J]. Crops, 2015(4): 152-156. |
| [38] |
王吉秀, 李想, 李祖然, 等. 小花南芥与玉米间作对Pb化学形态及富集特征的影响[J]. 环境工程学报, 2018, 12(12): 3456-3467. WANG Ji-xiu, LI Xiang, LI Zu-ran, et al. Effects of intercropping of Arabis alpina L.var. parviflora Franch and Zea mays L. on chemical speciation and accumulation characteristics of Pb[J]. Chinese Journal of Environmental Engineering, 2018, 12(12): 3456-3467. DOI:10.12030/j.cjee.201803151 |
| [39] |
王吉秀, 李元, 祖艳群, 等. 铅胁迫下小花南芥和玉米间作体系对植物生理的影响[J]. 环境科学与技术, 2017, 40(7): 54-59. WANG Ji-xiu, LI Yuan, ZU Yan-qun, et al. Effects of Arabis alpina L.var. parviflora Franch and Zea mays L. in an intercropping system on plant physiology to lead stress[J]. Environmental Science and Technology, 2017, 40(7): 54-59. |