 2019, Vol. 38
2019, Vol. 38



2. 农业农村部环境保护科研监测所, 天津 300191;
3. 贵州师范大学贵州省山地环境信息系统与生态环境保护重点实验室, 贵阳 550001
2. Agro-Environmental Protection Institute, Ministry of Agriculture and Rural Affairs, Tianjin 300191, China;
3. Guizhou Provincial Key Laboratory for Information System of Mountainous Areas and Protection of Ecological Environment, Guizhou Normal University, Guiyang 550001, China
Cd是八大土壤重金属污染元素之首,其污染范围广,污染程度深,对人体健康危害大,且不易转移、不易降解,导致Cd污染土壤的修复治理难度非常大。Cd污染来源可分为自然来源与人为来源两大类,自然因素包括成土母质、火山喷发以及岩石风化等,人为因素包括工业采矿、金属电镀及冶炼、化石燃料燃烧、大气沉降、汽车尾气排放、生活废水排放、农药化肥的不合理施用、污水灌溉等。《全国土壤污染状况调查报告》 [1]指出,我国土壤重金属污染严重,其中Cd点位超标率为7.0%,位列第一。据统计,我国受Cd污染的耕地面积已超过130 km2[2]。植物在Cd污染土壤中生长,其根系活性氧增加而受到氧化损伤[3]。植物受到Cd毒害导致生长受到抑制,同时Cd会被植物吸收积累,随着食物链进入人体内。Cd是人体非必需元素,当人体内Cd积累到一定量时,会导致高血压、骨质疏松、肝肾功能衰竭以及引起细胞癌变、突变等,Cd中毒可患“痛痛病”,使骨骼变形或导致骨折[4]。可见,Cd污染百害无一利,因而必须对Cd污染土壤进行修复治理。
硼酸(Orthoboric acid)作为硼(B)肥中常见的原料,可为植物生长提供B元素。B能促进植物体内碳水化合物的运输,有利于细胞壁的合成与稳定,并影响着蛋白质和核酸的代谢[5],还能缓解逆境胁迫对植物的毒害作用。褪黑素(Melatonin)是一种吲哚类激素,参与生物体内多种调节过程。在Cd胁迫下,添加褪黑素可提高水稻体内POD、SOD和CAT活性,降低MDA含量,促进种子萌发及幼苗生长[6],同时还能降低植物对Cd的吸收积累[7]。钼酸钠(Sodium molybdate dihydrate)通常作为钼(Mo)肥和复合肥料的成分之一,为植物生长提供养分。植物体内含多种Mo酶,能调节多种代谢过程[8],Mo还能提高植物抗寒[9]、抗干旱[10]、抗盐害[11]、抗重金属胁迫[12]的能力。硅酸钠(Sodium silicate)作为硅(Si)肥的原料之一,近年来在修复重金属污染土壤方面发挥着越来越重要的作用[13]。研究表明,添加硅酸钠能显著降低土壤有效态Cd含量以及水稻籽粒中的Cd含量[14]。
以上不同改良剂对植物重金属胁迫的缓解作用尽管已有一些研究,但其对Cd胁迫下水稻种子萌发、水稻吸收积累Cd的影响差异还未见报道。本文在同一生长条件下比较硼酸、褪黑素、钼酸钠和硅酸钠对水稻种子萌发、水稻吸收积累Cd的影响,以期筛选出有效治理Cd污染的改良剂,为Cd污染农田大面积治理推广应用提供科学依据。
1 材料与方法 1.1 试验材料 1.1.1 水稻品种供试水稻品种为潭两优83(国审稻2010002),记为T83,属籼型两系杂交水稻,品种来源于潭农S和潭早183,熟期适中,产量高。T83在长江中下游作双季早稻种植,全生育期平均109.4 d,株型适中,株高82.7 cm,穗长19.4 cm,每穗总粒数109.1粒,结实率84.4%,千粒重26.1 g(选育单位:湘潭市农业科学研究所)。
1.1.2 化学试剂及改良剂试验所用Cd为氯化镉,分析纯;改良剂硼酸、硅酸钠和钼酸钠均为分析纯;褪黑素为优级纯。
1.2 试验方案 1.2.1 萌发试验设置0、1、5 μmol·L-1 3个Cd浓度处理,5种改良剂处理:不添加改良剂(CK)、1 mg·L-1硼酸(B)、10 μmol·L-1褪黑素(MT)、0.5 mg·L-1钼酸钠(Mo)和1 mmol·L-1硅酸钠(Si),共15组处理,每组处理重复3次。硼酸、褪黑素、钼酸钠和硅酸钠处理浓度根据预实验以及笔者的前期研究报道设置[6, 12, 15-16]。选取适量籽粒饱满、颗粒大小相当的T83水稻种子,用5%的过氧化氢(H2O2)浸泡15 min以消毒,用去离子水反复冲洗3~5遍,将消毒洗净的T83种子放入28 ℃的恒温电热培育箱中避光催芽1 d。挑选露白状况大致相同的种子均匀摆放在铺有两层直径为90 mm滤纸的玻璃培养皿中,每个培养皿放30粒种子,根据试验设置加入5 mL不同Cd浓度处理液和5 mL不同改良剂处理液组合(CK用去离子水代替),共45个培养皿,放入28 ℃的恒温电热培育箱中避光萌发。培养期间每日更换一次滤纸及处理液,并定期观察记录种子的萌发情况。
1.2.2 水培试验水培试验与萌发试验中设置的Cd浓度和不同改良剂浓度均相同,即3个Cd浓度和5个改良剂处理,共15个处理,每个处理3次重复。选取籽粒饱满、颗粒大小相当的T83水稻种子若干,用5%的H2O2消毒浸泡15 min,用去离子水反复冲洗3~5遍。将种子均匀洒在塑料育苗盘上,种子的间距不宜过密,以确保种子萌发及幼苗生长有充足的氧气和生长空间,育苗盘中的去离子水以刚好浸没种子为宜。将盛有种子的育苗盘放入28 ℃的恒温电热培育箱中避光催芽萌发,1 d后将露白萌发的种子转移至人工气候室内(条件:温度为28 ℃、湿度为95%、光照为100%)培养,遮光处理2 d。一周后将水稻幼苗转移至容积为8 L的聚乙烯(PE)水培箱中继续培养,用高密度净化海绵包裹水稻幼苗进行固定,每个孔移栽4株,每株水稻幼苗间相隔一定的距离,以保证幼苗生长有足够的空间。用pH值为5.5、浓度1/8的Hoagland营养液(表 1)培养一周。待水稻幼苗长至三叶一心期时,将水培箱中的溶液更换为去离子水,对水稻幼苗进行饥饿处理1 d,再分别对幼苗进行不同的试验处理。
|
|
表 1 营养液配方(mg·L-1) Table 1 Nutrient solution formula (mg·L-1) |

挑选长势大致相同的水稻幼苗移至500 mL的PVC罐中(高14 cm、内口径7 cm),每罐移栽4株,每株水稻幼苗间隔一定距离,以保证幼苗生长有足够的空间。为了避免气候室每个位置光线强弱差异对水稻幼苗生长的影响,每日变换一次PVC罐放置的位置。PVC罐中液体为Hoagland营养液和不同Cd浓度处理液与不同改良剂处理液,处理液每3 d更换一次,并在第8 d和第22 d分别收取水稻样品。
1.3 测定方法 1.3.1 萌发试验指标测定萌发试验阶段,分别在添加处理液3 d和7 d后统计有效发芽的种子数,用于计算发芽势和发芽率。水稻种子根长大于种子长,且芽长比种子长一半大时算作有效发芽。

|
处理7 d后收取样品,用直尺测量水稻幼苗的根长和芽长,取平均值。将水稻样品拆分为根和芽,用天平称量水稻幼苗的根鲜质量和芽鲜质量,按每皿计算。分别将根和芽装入信封中,置于80 ℃的烘箱中烘干至恒质量,采用硝酸消解法进行消解,随后用原子吸收分光光度计-石墨炉测定水稻样品根系和幼芽中的Cd含量。
1.3.2 水培试验指标测定水培试验阶段,在添加处理液之后的第8 d和第22 d,从每个PVC罐中随机收取2株水稻苗,将其根部放在5%的氯化钙溶液中浸泡30 min,再用去离子水冲洗、浸泡,反复3次。将清洗好的水稻样品拆分为根和茎叶分别装入信封中,置于80 ℃的烘箱中烘干至恒质量,采用硝酸消解法进行消解,随后用原子吸收分光光度计-石墨炉测定水稻样品根系和茎叶中的Cd含量。
1.4 数据处理用Microsoft Excel 2010和SPSS 21.0进行试验数据分析以及数据图标绘制,并对不同处理间的数据用单因素方差分析(ANOVA)进行两两比较(Duncan多重比较)以检验差异显著性,试验数据用平均值±标准偏差表示。
2 结果与分析 2.1 不同改良剂对Cd胁迫下水稻种子萌发的影响 2.1.1 水稻种子发芽情况没有Cd胁迫时,添加4种改良剂对种子发芽势无显著影响,Cd胁迫后4种改良剂对水稻种子发芽势均有显著的促进作用(图 1)。1 μmol·L-1和5 μmol·L-1Cd胁迫下水稻种子发芽势与CK相比分别降低22.2%和31.1%。在1 μmol·L-1Cd胁迫下,添加硼酸、褪黑素、钼酸钠和硅酸钠使水稻种子发芽势分别比CK处理提高15.6%、16.7%、16.7%和17.8%;在5 μmol·L-1Cd胁迫下添加这4种改良剂使水稻种子发芽势分别提高17.8%、17.8%、17.8%和22.2%,4种改良剂缓解Cd胁迫下水稻种子发芽势的效果相近。

|
不同小写字母表示同一Cd浓度处理下添加不同改良剂处理间差异显著(P<0.05)。下同 Different lowercase letters indicate significant differences (P < 0.05) among the treatments with the same Cd concentration and different amendments. The same below 图 1 不同改良剂对Cd胁迫下水稻种子发芽势的影响 Figure 1 Effects of different amendments on germination potential of rice seeds under Cd stress |
图 2为Cd胁迫下不同改良剂对水稻种子发芽率的影响。如图所示,Cd胁迫对种子发芽率也有抑制作用。1 μmol·L-1和5 μmol·L-1Cd胁迫下水稻种子发芽率相比CK处理分别降低8.9%和16.7%。在1 μmol·L-1Cd胁迫下,添加硼酸、褪黑素、钼酸钠和硅酸钠使水稻种子发芽率分别比CK处理提高6.7%、7.8%、7.8%和5.6%,在5 μmol·L-1Cd胁迫下分别提高11.1%、10.0%、11.1%和10.0%。

|
图 2 不同改良剂对Cd胁迫下水稻种子发芽率的影响 Figure 2 Effects of different amendments on germination rate of rice seeds under Cd stress |
从表 2中可以看出,水稻幼根和幼芽生长均明显受到Cd胁迫的抑制,尤其是对根系生长的抑制作用更强。随着Cd浓度增加,水稻根长和芽长相对减少。与CK处理相比,1 μmol·L-1 Cd胁迫下水稻根长和芽长分别减少1.47 cm和0.62 cm。当Cd浓度为5 μmol·L-1时,水稻根长比CK处理减少3.01 cm,芽长减少0.93 cm。添加不同改良剂对Cd胁迫下水稻根、芽生长均有促进作用。当Cd浓度为1 μmol·L-1时,添加硼酸、褪黑素、钼酸钠和硅酸钠使水稻根长分别比CK提高10.2%、10.9%、12.0%和10.5%,芽长除了硅酸钠处理显著提高11.0%外,其他改良剂处理影响不显著。当Cd浓度为5 μmol·L-1时,添加硼酸、褪黑素、钼酸钠和硅酸钠使水稻幼根长分别比CK提高41.3%、40.9%、40.1%和37.5%,芽长除了硅酸钠处理显著提高12.6%外,其他改良剂处理影响不显著。4种改良剂中,硅酸钠促进水稻幼芽生长效果较好。
|
|
表 2 不同改良剂对Cd胁迫下水稻根长和芽长的影响 Table 2 Effects of different amendments on root and shoot length of rice under Cd stress |
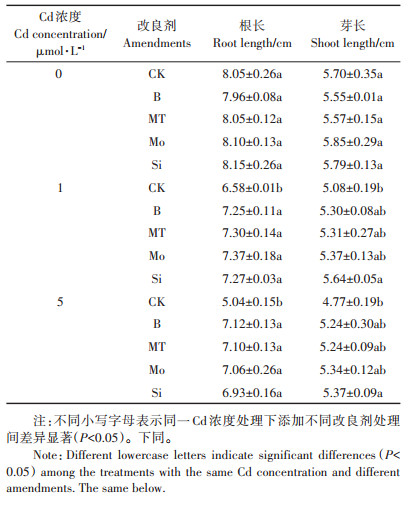
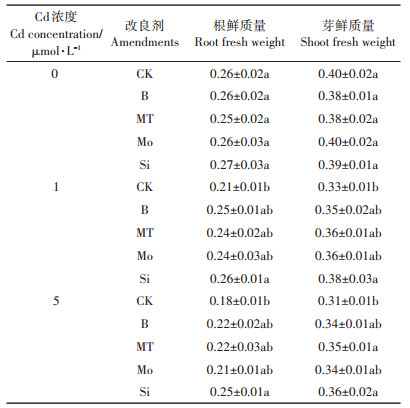
表 3为不同改良剂对Cd胁迫下水稻根鲜质量和芽鲜质量的影响,在Cd胁迫下,水稻根鲜质量和芽鲜质量均有所下降。Cd浓度为5 μmol·L-1时,水稻根鲜质量比CK处理降低0.08 g,幼芽鲜质量降低0.09 g。Cd浓度为1 μmol·L-1时,添加硅酸钠使水稻根鲜质量和芽鲜质量分别比CK显著提高23.8%和15.2%,添加其他改良剂处理影响效果不显著。Cd浓度为5 μmol·L-1时,添加硅酸钠后水稻根鲜质量和芽鲜质量分别比CK显著提高38.9%和16.1%,添加其他改良剂处理影响也不显著(MT处理的芽鲜质量除外)。
|
|
表 3 不同改良剂对Cd胁迫下水稻根鲜质量和芽鲜质量的影响(g) Table 3 Effects of different amendments on root and shoot fresh weight of rice under Cd stress (g) |
Cd胁迫下不同改良剂对水稻幼根中Cd含量的影响如图 3所示,可以看出不同改良剂均显著降低了水稻根系Cd含量(P<0.05)。当Cd浓度为1 μmol·L-1时,添加硼酸、褪黑素、钼酸钠和硅酸钠使水稻幼根Cd含量分别比CK处理降低53.1%、53.6%、54.6%和48.5%;当Cd浓度为5 μmol·L-1时,水稻幼根Cd含量分别降低50.1%、51.3%、47.0%和38.6%。

|
图 3 Cd胁迫下不同改良剂对种子萌发过程水稻幼根Cd含量的影响 Figure 3 Effects of different amendments on Cd concentrations of rice root during seed germination under Cd stress |
图 4为水稻幼芽中的Cd含量。当Cd浓度为1 μmol·L-1时,添加硼酸、褪黑素、钼酸钠和硅酸钠后水稻幼芽Cd含量分别比CK处理显著降低21.5%、24.6%、30.7%和23.2%;当Cd浓度为5 μmol·L-1时,水稻幼芽Cd含量分别显著降低36.9%、41.1%、39.3%和39.5%。

|
图 4 Cd胁迫下不同改良剂对种子萌发过程水稻幼芽Cd含量的影响 Figure 4 Effects of different amendments on Cd concentrations in shoot of rice during seed germination under Cd stress |
图 5为水培试验处理8 d后不同改良剂对Cd胁迫下水稻根系Cd含量的影响。当Cd浓度为1 μmol·L-1时,添加硼酸、褪黑素、钼酸钠和硅酸钠使水稻根系Cd含量分别比CK处理降低32.6%、38.3%、47.0%和72.5%(P<0.05);当Cd浓度为5 μmol·L-1时分别降低4.3%、21.0%、20.2%和23.3%。4种改良剂中,硅酸钠处理效果最明显。

|
图 5 水培试验中不同改良剂对Cd胁迫下水稻根系Cd含量的影响(8 d) Figure 5 Effects of different amendments on Cd concentrations of rice roots under Cd stress in solution culture (8 d) |
当Cd浓度为1 μmol·L-1时,添加硼酸、褪黑素和钼酸钠对水稻茎叶Cd含量影响不显著(P>0.05),但是硅酸钠处理使水稻茎叶Cd含量比CK显著降低75.1%(图 6);当Cd浓度为5 μmol·L-1时,添加硼酸对水稻茎叶Cd含量影响不显著,但是添加褪黑素、钼酸钠和硅酸钠使水稻茎叶Cd含量分别比CK处理显著降低31.9%、27.9%和63.1%,硅酸钠处理效果最显著,硼酸处理效果最差。

|
图 6 水培试验中不同改良剂对Cd胁迫下水稻茎叶Cd含量的影响(8 d) Figure 6 Effects of different amendments on Cd concentrations of rice stem and leaf under Cd stress in solution culture (8 d) |
图 7为水培处理22 d后不同改良剂对水稻根系Cd含量的影响。当Cd浓度为1 μmol·L-1时,添加硼酸对水稻根系Cd含量影响不显著(P>0.05),但是褪黑素、钼酸钠和硅酸钠处理使水稻根系Cd含量分别比CK显著降低17.7%、25.7%和33.7%;当Cd浓度为5 μmol·L-1时,添加硼酸、褪黑素、钼酸钠和硅酸钠使水稻根系Cd含量分别比CK处理显著降低6.7%、13.8%、13.1%和14.4%。对于水稻茎叶而言(图 8),当Cd浓度为1 μmol·L-1时,添加硼酸和褪黑素对水稻茎叶Cd含量影响不显著(P>0.05),但是钼酸钠和硅酸钠处理使水稻茎叶Cd含量分别比CK显著降低44.2%和41.5%;当Cd浓度为5 μmol·L-1时,硼酸和钼酸钠降低Cd含量效果不明显,但是褪黑素和硅酸钠使水稻茎叶Cd含量分别比CK显著降低33.1%和50.5%(P<0.05)。

|
图 7 水培试验中不同改良剂对Cd胁迫下水稻根系Cd含量的影响(22 d) Figure 7 Effects of different amendments on Cd concentrations of rice root under Cd stress in solution culture (22 d) |

|
图 8 水培试验中不同改良剂对Cd胁迫下水稻茎叶Cd含量的影响(22 d) Figure 8 Effects of different amendments on Cd concentrations of rice stem and leaf under Cd stress in solution culture (22 d) |
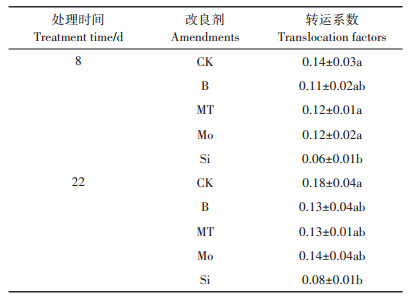
表 4为不同改良剂对5 μmol·L-1Cd胁迫下水稻根系到茎叶的转运系数的影响。与没有添加任何改良剂的CK处理相比,添加4种改良剂均能降低水稻的转运系数,但是除了硅酸钠处理达到显著水平(P<0.05)外,其余3种改良剂均不存在显著差异。从表 4中可以看出,随着培养时间的增加,Cd从水稻根系到茎叶的转运系数在增大,这种现象在不同改良剂处理下保持一致。从处理8 d到处理22 d,不添加改良剂时水稻转运系数提高了0.04,添加硼酸、褪黑素、钼酸钠和硅酸钠处理时水稻转运系数分别增加了0.02、0.01、0.02和0.02。
|
|
表 4 水培试验中不同改良剂对5 μmol·L-1Cd胁迫下水稻转运系数的影响 Table 4 Effects of different amendments on translocation factors of rice under 5 μmol·L-1 Cd stress in solution culture |
水稻是我国最主要的粮食作物,尤其在南方地区普遍种植水稻,而我国南方地区农田Cd污染问题比较严重,这不仅影响水稻生长与稻米产量,更会危害人体健康。Cd对水稻萌发及生长有抑制作用,且不同水稻品种对Cd的耐受性有所不同[17]。陈京都等[18]发现,Cd胁迫不仅影响水稻产量,还会抑制水稻对矿质营养元素Mg、P和K的吸收。本研究结果表明,Cd胁迫下水稻种子发芽势和发芽率显著降低,水稻根长、芽长、根鲜质量和芽鲜质量均明显减少。随着水稻培养时间的延长,水稻根系向茎叶转运的Cd会逐渐增多,表明水稻品种T83具有较强的Cd转运能力,这与胡莹等[19]的研究中籼稻品种对Cd的转运情况一致。由于水稻籽粒Cd含量与根系向茎叶转运Cd存在显著正相关关系[20],因此可推测T83可能存在水稻籽粒Cd含量较高的风险。
添加1 mg·L-1硼酸处理使水稻种子发芽势和发芽率、水稻根长显著提高,降低种子萌发过程中水稻幼根和幼芽Cd含量,说明B肥对水稻Cd污染防治有一定作用,与前人的研究结果相一致[21]。植物吸收的B大部分靠土壤提供,而我国许多省份耕层土壤中B元素含量不足,土壤中的B可能随淋溶过程流失,导致植物生长过程中B素缺乏,因此B肥的施用尤为重要。对于重金属污染农田土壤而言,适量施加B肥不仅能提高作物的产量,还能缓解重金属对植物的胁迫作用,降低作物食用部分的重金属含量。研究表明,B可通过络合膜组分来稳定质膜结构,与酚类物质结合从而保护细胞膜免受毒害,这可能是B抗逆境胁迫的机制之一[22]。拉飞克等[21]研究表明,添加0.5 mg·L-1的B能显著减缓Cd胁迫下植物POD、SOD和CAT活性的下降,降低O2-·产生速率,使植物的抗氧化能力得到提高,同时植物体内叶绿素和可溶性蛋白含量显著增加,从而提高了槐叶萍抵抗Cd胁迫的能力。叶绿素影响着植物的光合作用,可溶性蛋白能调节水平衡,可溶性糖能调节渗透性,脯氨酸可对细胞膜和蛋白质起到保护作用,并参与活性氧自由基的清除,这些物质对植物抵抗重金属的迫害都有一定的帮助,这也可能是B有抗逆性的原因之一。还有可能是B与重金属之间存在拮抗作用和竞争吸收,因而添加B能够减少水稻对重金属的吸收[15]。
本研究中,在5 μmol· L-1Cd胁迫下,添加10 μmol·L-1褪黑素处理使水稻种子发芽势和发芽率分别提高17.8%和10%,种子萌发过程中水稻幼根和幼芽Cd含量分别降低51.3%和41.1%,说明褪黑素缓解Cd污染胁迫下水稻生长的效果显著,这与刘仕翔等[6]的研究结果一致。褪黑素是目前发现的抗氧化作用最强的自由基清除剂,能有效缓解Cd对植物胚芽和生殖组织的氧化损伤[23]。在萌发试验中,褪黑素促进了水稻根芽生长,根系作为水稻吸收营养物质的重要器官,直接反映了Cd毒害作用对水稻的影响。Sarropoulou等[23]证明,褪黑素确实具有促进植物根系生长的作用。Posmyk等[24]研究表明,在Cu2+胁迫条件下,经褪黑素预处理后的红花甘蓝种子发芽率显著提高。黄佳璟等[7]发现,喷施褪黑素能够提高Cd胁迫下的萝卜生物量、抗氧化酶活性、可溶性蛋白含量、叶绿素含量等,同时,还能降低植物中的Cd含量,促进萝卜生长。唐懿等[25]研究发现,褪黑素浸种对植物有生长调节作用,能促进豌豆幼苗生长,提高叶绿素含量,增强光合作用,缓解了Cd对豌豆幼苗的毒害。褪黑素缓解重金属胁迫的机制可能是因为褪黑素能够和金属离子进行螯合,形成毒性较低或无毒的物质从而降低重金属对植物生长的毒害[26]。
添加0.5 mg·L-1钼酸钠处理对Cd胁迫下水稻种子发芽势和发芽率也有显著提高,种子萌发和水培实验中水稻植株Cd含量也显著降低(图 3~图 5),证实钼酸钠也是一种缓解水稻Cd污染胁迫的有效改良剂。Mo是植物和动物正常生长必不可缺的营养元素之一,植物补充Mo元素能够防治叶片卷曲、枯黄等病害[26]。Babenko等[27]通过种子预处理和喷施叶面肥来激活植物中含Mo酶的活性,提高了哈萨克斯坦北部草原区牧草的产量,该研究发现,Mo处理可以增加硝酸还原酶、醛氧化酶和黄嘌呤脱氢酶的活性,从而降低植物组织的氧化损伤,改善植物健康。孙学成等[28]研究表明,施Mo能有效提高植物叶片抗氧化酶活性,提高植物抗寒能力。聂呈荣等[29]表示,经过钼酸钠浸种后花生种子发芽率得到提高,有效促进了种子胚根的生长,减缓了清除活性氧能力的下降,维持了较好的自我保护能力。Mo肥对降低蔬菜中的硝酸盐含量也有明显的作用[30]。袁彪等[12]发现,添加Mo能显著降低As胁迫下水稻根系及茎叶的As积累,增加生物量,促进水稻生长。本研究中,添加钼酸钠显著促进了水稻生长发育,有效缓解了Cd对植物的毒害,其原理可能是因为植物中有多种酶含Mo,施Mo后能影响含Mo酶的活性而使其发挥作用。还有可能是因为Mo是高等植物生长的必需元素,它能促进植物生长发育,从而增加植物的抗逆性。
本研究发现,Cd胁迫下添加1 mmol·L-1硅酸钠处理均使水稻种子发芽势、发芽率、根长、芽长、根鲜质量、芽鲜质量比CK处理显著提高;种子萌发和水培实验中水稻植株Cd含量也显著降低,而且4种改良剂中硅酸钠处理的效果最显著,它是一种缓解水稻Cd污染胁迫的最有效的改良剂。已经有大量的研究结果从促进植物生长和降低重金属迁移性等多方面证实了Si对缓解重金属胁迫有显著效果,这与本试验研究结果相一致。孙岩等[31]研究表明,Si能降低水稻中Cd的迁移性,从而缓解Cd对水稻的毒害作用。黄秋婵等[32]研究表明,加Si能够促进水稻吸收Mg、Cu、Zn、Fe元素以提高光合作用产物。葛永红等[33]用硅酸钠溶液对李杏果实进行浸泡处理,研究结果表明,硅酸钠可以提高活性氧的产生,调节酶活性以及苯丙烷代谢,增加果实对病害的抗性。薛高峰等[34]首次发现了施Si能提高由细菌引起的水稻抗白叶枯病害的能力,同时能显著增加水稻植株的生物量。在土壤重金属修复中,硅酸钠是常见的钝化剂,主要修复机理是提高土壤pH值、改变重金属在土壤中的存在形态等[35]。Chen等[36]研究表明,水稻叶面喷施纳米Si能显著降低籽粒中Cd的积累。向猛等[16]研究表明,加Si能明显降低Sb(Ⅲ)和Sb(Ⅴ)在水稻根系及茎叶中的积累量,说明Si能有效缓解重金属对水稻的毒害作用。王怡璇等[14]发现,施Si不仅能增加水稻产量,还能促进水稻根表铁膜的形成,从而降低Cd的转运能力,减少籽粒中Cd的含量。史新慧等[37]通过对Si结合蛋白的定位表示,抑制质外体途径的运输量从而降低了Cd向茎叶的运输。
4 结论(1)萌发试验中,4种改良剂硼酸、褪黑素、钼酸钠和硅酸钠均显著提高Cd胁迫下水稻种子的发芽势和发芽率,促进水稻种子根芽生长,降低水稻对Cd的吸收积累,对水稻Cd毒害起到了一定的缓解作用。
(2)水培实验中,4种改良剂也能不同程度地降低水稻根系和茎叶中的Cd含量,其中硅酸钠的处理效果最明显,其次是钼酸钠和褪黑素,硼酸的效果稍差。本研究结果显示,硅酸钠能更有效地缓解Cd毒害,下一步可在Cd污染农田进行试验验证,从而在农田Cd污染治理中推广应用。
| [1] |
环境保护部, 国土资源部.全国土壤污染状况调查公报[R].北京: 环境保护部, 国土资源部, 2014. Ministry of Environmental Protection, Ministry of Land and Resources. National survey of soil pollution[R]. Beijing: Ministry of Environmental Protection, Ministry of Land and Resources, 2014. http://www.cnki.com.cn/Article/CJFDTotal-ZHBY201405004.htm |
| [2] |
吴燕玉, 周启星, 田均良. 制定我国土壤环境标准(汞、镉、铅和砷)的探讨[J]. 应用生态学报, 1991, 2(4): 334-339. WU Yan-yu, ZHOU Qi-xing, TIAN Jun-liang. An approach to enactment of soil-environmental standards (Hg, Cd, Pb and As) in China[J]. Chinese Journal of Applied Ecology, 1991, 2(4): 334-339. |
| [3] |
肖清铁, 王经源, 郑新宇, 等. 水稻根系响应镉胁迫的蛋白质差异表达[J]. 生态学报, 2015, 35(24): 8276-8283. XIAO Qing-tie, WANG Jing-yuan, ZHENG Xin-yu, et al. Analysis of the differently expressed proteins in rice roots in response to cadmium stress[J]. Acta Ecologica Sinica, 2015, 35(24): 8276-8283. |
| [4] |
黄益宗, 郝晓伟, 雷鸣, 等. 重金属污染土壤修复技术及其修复实践[J]. 农业环境科学学报, 2013, 32(3): 409-417. HUANG Yi-zong, HAO Xiao-wei, LEI Ming, et al. The remediation technology and remediation practice of heavy metals-contaminated soil[J]. Journal of Agro-Environment Science, 2013, 32(3): 409-417. |
| [5] |
O'Neill M A, Ishii T, Albersheim P, et al. Rhamnogal acturonan Ⅱ:Structure and function of a borate cross-linked cell wall pectic polysaccharide[J]. Annual Review of Plant Biology, 2004, 55(1): 109-139. DOI:10.1146/annurev.arplant.55.031903.141750 |
| [6] |
刘仕翔, 黄益宗, 罗泽娇, 等. 外源褪黑素处理对镉胁迫下水稻种子萌发的影响[J]. 农业环境科学学报, 2016, 35(6): 1034-1041. LIU Shi-xiang, HUANG Yi-zong, LUO Ze-jiao, et al. Effects of exogenous melatonin on germination of rice seeds under Cd stresses[J]. Journal of Agro-Environment Science, 2016, 35(6): 1034-1041. |
| [7] |
黄佳璟, 林立金, 陈发波, 等. 喷施褪黑素对萝卜生长及镉积累的影响[J]. 四川农业大学学报, 2017, 35(3): 375-380. HUANG Jia-jing, LIN Li-jin, CHEN Fa-bo, et al. Effects of spraying melatonin on growth and cadmium accumulation of radish[J]. Journal of Sichuan Agricultural University, 2017, 35(3): 375-380. |
| [8] |
Mendel R R. Metabolism of molybdenum[J]. Metal Ions in Life Sciences, 2013, 12: 503-528. DOI:10.1007/978-94-007-5561-1 |
| [9] |
武松伟, 胡承孝, 谭启玲, 等. 钼与植物抗寒性研究进展[J]. 湖北农业科学, 2016, 55(1): 13-16. WU Song-wei, HU Cheng-xiao, TAN Qi-ling, et al. Advances in molybdenum nutrition and cold resistance in plant[J]. Hubei Agricultural Sciences, 2016, 55(1): 13-16. |
| [10] |
Wu S, Hu C, Tan Q, et al. Effects of molybdenum on water utilization, antioxidative defense system and osmotic-adjustment ability in winter wheat (Triticum aestivum) under drought stress[J]. Plant Physiology and Biochemistry, 2014, 83: 365-374. DOI:10.1016/j.plaphy.2014.08.022 |
| [11] |
Zhang M, Hu C X, Zhao X H, et al. Molybdenum improves antioxidant and osmotic-adjustment ability against salt stress in Chinese cabbage (Brassica campestris L. ssp. Pekinensis)[J]. Plant and Soil, 2012, 355(1/2): 375-383. |
| [12] |
袁彪, 黄益宗, 蔡立群, 等. 外源Mo降低As(Ⅲ)和As(Ⅴ)对水稻的毒性及As的积累[J]. 生态毒理学报, 2017, 12(1): 219-227. YUAN Biao, HUANG Yi-zong, CAI Li-qun, et al. Molybdenum reduced the toxicity and accumulation of As(Ⅲ) and As(Ⅴ) in rice seedling[J]. Asian Journal of Ecotoxicology, 2017, 12(1): 219-227. |
| [13] |
孙岩, 韩颖, 李军, 等. 硅对镉胁迫下水稻生物量及镉的化学形态的影响[J]. 西南农业学报, 2013, 26(3): 1240-1244. SUN Yan, HAN Ying, LI Jun, et al. Effect of Si on rice biomass and chemical species of Cd under Cd stress[J]. Southwest China Journal of Agricultural Sciences, 2013, 26(3): 1240-1244. DOI:10.3969/j.issn.1001-4829.2013.03.078 |
| [14] |
王怡璇, 刘杰, 唐云舒, 等. 硅对水稻镉转运的抑制效应研究[J]. 生态环境学报, 2016, 25(11): 1822-1827. WANG Yi-xuan, LIU Jie, TANG Yun-shu, et al. Inhibitory effect of silicon on cadmium accumulation and transportation in rice[J]. Ecology and Environmental Sciences, 2016, 25(11): 1822-1827. |
| [15] |
向猛, 黄益宗, 蔡立群, 等. 硼锑交互作用对水稻吸收积累锑和硼的影响[J]. 环境科学, 2015, 36(4): 1474-1480. XIANG Meng, HUANG Yi-zong, CAI Li-qun, et al. Effect of boronantimony interaction on the uptake and accumulation of antimony and boron by rice seedling[J]. Environmental Science, 2015, 36(4): 1474-1480. |
| [16] |
向猛, 黄益宗, 蔡立群, 等. 水稻吸收积累硅和锑的相互影响水培试验研究[J]. 农业环境科学学报, 2014, 33(11): 2090-2097. XIANG Meng, HUANG Yi -zong, CAI Li-qun, et al. Interactive effects of antimony and silicon on their uptake and accumulation by rice seedling in solution culture[J]. Journal of Agro-Environment Science, 2014, 33(11): 2090-2097. DOI:10.11654/jaes.2014.11.003 |
| [17] |
孙亚莉, 刘红梅, 徐庆国. 镉胁迫对不同水稻品种种子萌发特性的影响[J]. 中国水稻科学, 2017, 31(4): 425-431. SUN Ya-li, LIU Hong-mei, XU Qing-guo. Effects of cadmium stress on rice seed germination characteristics[J]. Chinese Journal of Rice Science, 2017, 31(4): 425-431. |
| [18] |
陈京都, 何理, 许轲, 等. 镉胁迫对不同基因型水稻生长及矿质营养元素吸收的影响[J]. 生态学杂志, 2013, 32(12): 3219-3225. CHEN Jing-du, HE Li, XU Ke, et al. Growth and nutritional element absorption of different rice genotypes under cadmium stress[J]. Chinese Journal of Ecology, 2013, 32(12): 3219-3225. |
| [19] |
胡莹, 黄益宗, 段桂兰, 等. 镉对不同生态型水稻的毒性及其在水稻体内迁移转运[J]. 生态毒理学报, 2012, 7(6): 664-670. HU Ying, HUANG Yi-zong, DUAN Gui-lan, et al. Cadmium toxicity and its translocation in two ecotype rice cultivars[J]. Asian Journal of Ecotoxicology, 2012, 7(6): 664-670. |
| [20] |
冯莲莲, 郭京霞, 黄梓璨, 等. 水稻土中7个水稻品种对土壤Cd、Pb的富集与转运:田间研究[J]. 生态环境学报, 2017, 26(12): 2146-2153. FENG Lian-lian, GUO Jing-xia, HUANG Zi-can, et al. A field study on the accumulation and translocation of cadmium and lead from a contaminated paddy rice field by seven rice cultivars[J]. Ecology and Environmental Sciences, 2017, 26(12): 2146-2153. |
| [21] |
拉飞克, 施国新, 王学, 等. 硼对镉胁迫下槐叶萍生理生化特性的影响[J]. 南京师大学报(自然科学版), 2003, 26(3): 72-75. Khaled R, SHI Guo-xin, WANG Xue, et al. The effects of boron on the physiological properties of Salvinia natans(L.)leaves under the stress of the Cd[J]. Journal of Nanjing Normal University (Natural Science), 2003, 26(3): 72-75. DOI:10.3969/j.issn.1001-4616.2003.03.016 |
| [22] |
Manchester L C, Tan D X, Reiter R J, et al. High levels of melatonin in the seeds of edible plants:Possible function in germ tissue protection[J]. Life Sciences, 2000, 67(25): 3023-3029. DOI:10.1016/S0024-3205(00)00896-1 |
| [23] |
Sarropoulou V, Dimassi-Theriou K, Therios I, et al. Melatonin enhances root regeneration, photosynthetic pigments, biomass, total carbohydrates and proline content in the cherry rootstock PHL-C (Prunus avium×Prunus cerasus)[J]. Plant Physiology and Biochemistry, 2012, 61: 162-168. DOI:10.1016/j.plaphy.2012.10.001 |
| [24] |
Posmyk M M, Kuran H, Marciniak K, et al. Presowing seed treatment with melatonin protects red cabbage seedlings against toxic copper ion concentrations[J]. Journal of Pineal Research, 2008, 45(1): 24-31. DOI:10.1111/jpi.2008.45.issue-1 |
| [25] |
唐懿, 任纬, 刘副刚, 等. 褪黑素浸种对豌豆幼苗生长及镉积累的影响[J]. 土壤, 2018, 50(1): 109-114. TANG Yi, REN Wei, LIU Fu-gang, et al. Effects of melatonin soaking on growth and cadmium accumulation of pea seedlings[J]. Soils, 2018, 50(1): 109-114. |
| [26] |
曹恭, 梁鸣早. 钼:平衡栽培体系中植物必需的微量元素[J]. 土壤肥料, 2004(3): 52-55. CAO Gong, LIANG Ming-zao. Trace elements necessary for plants in molybdenum -balanced cultivation system[J]. Soils and Fertilizers, 2004(3): 52-55. |
| [27] |
Babenko O N, Brychkova G, Sagi M, et al. Molybdenum application enhances adaptation of crested wheat grass to salinity stress[J]. Acta Physiol Plant, 2015, 37(2): 1-13. DOI:10.1007/s11738-014-1746-y |
| [28] |
孙学成, 谭启玲, 胡承孝, 等. 低温胁迫下钼对冬小麦抗氧化酶活性的影响[J]. 中国农业科学, 2006, 39(5): 952-959. SUN Xue-cheng, TAN Qi-ling, HU Cheng-xiao, et al. Effects of molybdenum on antioxidative enzymes in winter wheat under low temperature stress[J]. Scientia Agricultura Sinica, 2006, 39(5): 952-959. DOI:10.3321/j.issn:0578-1752.2006.05.014 |
| [29] |
聂呈荣, 谢惠银, 温玉辉, 等. 钼酸钠浸种对花生种子萌芽及幼苗生长的影响[J]. 土壤肥料, 2003(4): 29-31. NIE Cheng-rong, XIE Hui-yin, WEN Yu-hui, et al. Effect of soaking seeds with natrium molybdate on seed germination and seedlings growth of peanut[J]. Soils and Fertilizers, 2003(4): 29-31. |
| [30] |
赵静, 白清云, 戴晓华, 等. 钼对降低蔬菜硝酸盐积累的效应研究[J]. 农业环境科学学报, 2001, 20(4): 238-239. ZHAO Jing, BAI Qing-yun, DAI Xiao-hua, et al. Alleviation of nitrate accumulation in vegetables by application of molybdenum[J]. Journal of Agro-Environment Science, 2001, 20(4): 238-239. DOI:10.3321/j.issn:1672-2043.2001.04.014 |
| [31] |
孙岩, 韩颖, 李军, 等. 硅对镉胁迫下水稻生物量及镉的化学形态的影响[J]. 西南农业学报, 2013, 26(3): 1240-1244. SUN Yan, HAN Ying, LI Jun, et al. Effect of Si on rice biomass and chemical species of Cd under Cd stress[J]. Southwest China Journal of Agricultural Sciences, 2013, 26(3): 1240-1244. DOI:10.3969/j.issn.1001-4829.2013.03.078 |
| [32] |
黄秋婵, 许元明, 曾振芳, 等. 硅对镉胁迫下水稻幼苗茎叶元素含量的影响[J]. 湖北农业科学, 2013, 52(11): 2489-2495. HUANG Qiu-chan, XU Yuan-ming, ZENG Zhen-fang, et al. Effect of silicon on element content in stems and leaves of seedlings in Oryza sativa under cadmium stress[J]. Hubei Agricultural Sciences, 2013, 52(11): 2489-2495. DOI:10.3969/j.issn.0439-8114.2013.11.003 |
| [33] |
葛永红, 李灿婴, 王毅, 等. 硅酸钠处理对杏果实活性氧和苯丙烷代谢的影响[J]. 食品工业科技, 2014, 35(13): 317-326. GE Yong-hong, LI Can-ying, WANG Yi, et al. Effects of sodium silicate dipping on reactive oxygen species metabolism and phenylpropanoid pathway in apricot fruit[J]. Science and Technology of Food Industry, 2014, 35(13): 317-326. |
| [34] |
薛高峰, 孙万春, 宋阿琳, 等. 硅对水稻生长、白叶枯病抗性及病程相关蛋白活性的影响[J]. 中国农业科学, 2010, 43(4): 690-697. XUE Gao-feng, SUN Wan-chun, SONG A-lin, et al. Influence of silicon on rice growth, resistance to bacterial blight and activity of pathogenesis-related proteins[J]. Scientia Agricultura Sinica, 2010, 43(4): 690-697. DOI:10.3864/j.issn.0578-1752.2010.04.004 |
| [35] |
赵明柳, 唐守寅, 董海霞, 等. 硅酸钠对重金属污染土壤性质和水稻吸收Cd、Pb和Zn的影响[J]. 农业环境科学学报, 2016, 35(9): 1653-1659. ZHAO Ming-liu, TANG Shou-yin, DONG Hai-xia, et al. Effects of sodium silicate on soil properties and Cd, Pb and Zn absorption by rice plant[J]. Journal of Agro-Environment Science, 2016, 35(9): 1653-1659. |
| [36] |
Chen R, Zhang C B, Zhao Y L, et al. Foliar application with nano-silicon reduced cadmium accumulation in grains by inhibiting cadmium translocation in rice plants[J]. Environmental Science and Pollution Research, 2018, 25(3): 2361-2368. DOI:10.1007/s11356-017-0681-z |
| [37] |
史新慧, 王贺, 张福锁. 硅提高水稻抗镉毒害机制的研究[J]. 农业环境科学学报, 2006, 25(5): 1112-1116. SHI Xin-hui, WANG He, ZHANG Fu-suo. Research on the mechanism of silica improving the resistance of rice seedlings to Cd[J]. Journal of Agro-Environment Science, 2006, 25(5): 1112-1116. DOI:10.3321/j.issn:1672-2043.2006.05.004 |