 2020, Vol. 39
2020, Vol. 39



2. 中国科学院大学, 北京 100049;
3. 山东潍坊烟草有限公司, 山东 潍坊 261205;
4. 南京师范大学地理科学学院, 江苏省地理信息资源开发与利用协同创新中心, 南京 210023
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Shandong Weifang Tobacco Co., Ltd., Weifang 261205, China;
4. School of Geography Sciences, Nanjing Normal University, Jiangsu Center for Collaborative Innovation in Geographical Information Resource Development and Application, Nanjing 210023, China
我国是农业大国,烟草作为我国的重要经济作物,烟叶生产量及成品烟销售量已占世界的1/3左右。根据2012年烟草专卖局官方统计,我国烟叶种植面积已达到3300万hm2。而与先进烤烟生产国相比较(如津巴布韦和美国),我国烟叶的内外在品质仍存在差距,表现在整体香气量不足,上部烟叶烟碱含量较高和叶片过厚等方面[1]。在烤烟生长发育所必需的营养元素中,氮素是影响烟叶产量和品质最为重要的营养元素[2],氮素不足或过量都会导致烟叶产量和品质的下降[3]。巨晓棠等[4]研究表明,我国烤烟在打顶后仍吸收大量氮素,且这部分氮素主要来自土壤氮,这与国外生产优质烤烟的吸氮曲线有很大差异。有研究表明,我国80%以上的植烟土壤中有机质含量高于25 g·kg-1[5],这意味着土壤氮在合适条件下会释放大量氮素,尤其在烤烟生长发育后期多是在高温多雨的气候条件下。另外,李春俭等[1]研究也表明,我国部分植烟区的烤烟在打顶后仍大量吸收氮素,是导致我国上部烟叶整体质量得不到提升的重要原因。因此,通过采取合适的措施来调控烤烟生长后期土壤氮素供应是获得优质烤烟烟叶的关键[6-7]。
硝态氮和铵态氮是烤烟能够利用的主要有效氮源,施入土壤中的氮素受微生物作用进行转化分配,其转化程度除受土壤质地、温度、水分、pH、有机质和含氮量等诸多因子的影响外[8-9],还受外源物质尤其是有机质的影响,这是由于有机物质可向微生物提供可利用的碳源和能源[10]。向土壤中添加不同来源和不同质量的外源碳对土壤氮素转化具有重要影响[11-13],所得出的结果由于添加碳源的性质不同及土壤本身的差异而有所不同,表现为促进或抑制了土壤氮的净矿化[14-16]。添加外源有机质后土壤氮素有效性取决于其C/N,添加低C/N的有机质(C/N < 20)表现为氮素的净矿化[17];而添加高C/N的有机质则导致微生物对氮素的净固持[18]。因此,微生物同化作用在调节土壤中有效氮含量方面起着重要的作用[19]。已有研究报道,异养微生物能够被可利用的碳源刺激而促进微生物同化作用[20-21]。几项研究表明,蔗糖、锯末或稻草的施用均能降低草地、草原和山艾树生态系统恢复时的土壤氮素有效性[22-23]。巨晓棠等[24]研究也发现,添加小麦秸秆(高C/N)显著降低了土壤中无机氮含量;而添加苜蓿秸秆或鸡粪(低C/N)则显著提高土壤中无机氮含量。
另有研究表明,添加外源碳影响了微生物对不同形态氮的吸收同化,因此对不同肥力水平土壤的氮素转化产生显著影响[12-13]。例如,长期受微生物优先利用NH4+-N这一传统观点的影响,人们普遍认为农田土壤微生物不利用NO3--N[25-26];但向土壤中添加作物残茬和收获后残留物,由于增加了土壤中有效碳含量,NO3--N的微生物同化量得到提高[27-28]。在一项Meta分析中,Cheng等[29]也发现当葡萄糖和醋酸盐的添加量≥500 mg C·kg-1土或者动物粪肥和植物秸秆的C/N> 18时,均会促进微生物对NO3--N的同化。
添加外源碳会影响土壤氮素的矿化作用和硝化作用,同时也会对反硝化过程产生影响[30]。N2O主要来自土壤中氮素硝化和反硝化两个过程。由于土壤中好气和厌气微区同时存在,因此两个过程可能同时发生。研究发现反应底物(氮源、碳源)的有效性是影响硝化和反硝化作用的重要因素之一。添加外源碳对土壤有机碳的转化过程产生重要影响,进而直接影响到土壤的碳固定和温室气体排放[31]。另外,Morley等[32]研究表明,土壤有机质为绝大多数异养微生物提供了碳源和电子供体,并为土壤呼吸作用提供基质。因此,土壤有机质是调控N2O排放的重要因子。另有研究表明,添加碳源能够增强土壤呼吸强度并显著增加土壤微生物量,但不同外源碳成分及浓度导致温室气体排放的多少也有差异[33-34]。
腐植酸是土壤有机质的主要成分,它是由动植物残体经化学反应和微生物分解形成的大分子有机酸[35]。腐植酸无毒、无公害,在农业生产中具有改良土壤、增进肥效、刺激生长、促进抗逆、改善品质等5大作用[36]。黄腐酸钾作为腐植酸类肥料的一种,具有短碳链分子结构和可溶性较强的特点。研究表明,氮肥配施黄腐酸钾均能提高作物产量和肥料利用率[37-38]。然而,黄腐酸钾作为一种高C/N的可溶性碳源,向土壤中添加黄腐酸钾对于氮素转化的影响如何?另外,在烤烟生长后期向土壤中添加黄腐酸钾是否可以作为一种调控植烟土壤后期氮素供应过多的措施?这些问题值得我们探究。因此本文拟先采用室内培养的实验方法,通过添加硝酸铵模拟植烟土壤后期含有大量有效氮的情景,探究添加不同量黄腐酸钾对植烟土壤氮素转化及N2O排放的影响,为下一步田间工作提供数据支撑。这对明确添加黄腐酸钾后氮素的转化规律具有重要意义,同时也可为烤烟或其他作物生产中合理施用黄腐酸钾提供科学依据。
1 材料与方法 1.1 供试土壤与材料供试土壤取自山东沂水县烟草实验站(118.63° E,35.85°N)内烟田0~20 cm表层。实验站位于鲁中南丘陵区,海拔191 m,属暖温带大陆性季风气候,年均气温14.1 ℃,降水量849 mm,蒸发量约为560 mm,无霜期200 d以上,土壤类型为褐土(简育干润雏形土)。供试土壤的基本理化性质为:可溶性全氮18.48 mg·kg-1,硝态氮7.58 mg·kg-1,铵态氮4.68 mg· kg-1,可溶性有机氮6.22 mg·kg-1,全氮0.76 g·kg-1,有机碳5.10 g·kg-1,C/N 11.55,pH 6.94。供试土壤多点采集混合后,挑出肉眼可见的植物残体和石块,过2 mm筛后保存于4 ℃冰箱。
黄腐酸钾由上海芳甸生物科技有限公司提供(全氮2.04 g·kg-1,全碳440.71 g·kg-1,以干基计),C / N 218.08。分别快速称取4.17、8.33、16.77 g和33.54 g黄腐酸钾溶于250 mL去离子水中,然后在超声波振荡器内振荡30 min,使其溶解均匀,配成4种不同浓度的黄腐酸钾溶液:0.83,1.67,3.33 g·mL-1和5 g·mL-1。
1.2 实验设计试验共设置5个处理:(1)对照(CK):只添加硝酸铵(含NH4+-N和NO3--N各100 mg·kg-1,下同);(2)T1:硝酸铵+2.5 g·kg-1黄腐酸钾;(3)T2:硝酸铵+5 g·kg-1黄腐酸钾;(4)T3:硝酸铵+10 g·kg-1黄腐酸钾;(5)T4:硝酸铵+15 g·kg-1黄腐酸钾。称取相当于20 g干土质量的新鲜土样于250 mL玻璃瓶中,培养开始前把土壤混匀,使其厚度均匀地平铺在瓶底,放在25 ℃室内预培养1 d。预培养结束后,用移液管向每个玻璃瓶中添加3 mL不同浓度的黄腐酸钾溶液,使黄腐酸钾添加量分别为2.5、5、10 g·kg-1和15 g·kg-1,CK处理则添加3 mL去离子水;同时向每个玻璃瓶中加3 mL NH4NO3溶液(200 mg·kg-1),含NH4+-N和NO3--N各100 mg·kg-1。所有添加溶液均用移液管均匀滴入,使其尽可能均匀分布于土壤中,此时土壤含水量为最大田间持水量的67%。然后用硅胶塞将玻璃瓶密封,继续在25 ℃下恒温培养30 d,每隔2 d去塞通气0.5 h左右,每2~3 d补水1次以补充因蒸发导致的水分损失。分别在添加氮肥和黄腐酸钾后第1、3、5、7、14、30 d采集气体样品,采集气体样品之前,需要提前6 h进行换气密封。换气前用南京大学研制的704硅橡胶将塞与瓶口缝隙密封,待硅胶干燥后,用真空泵抽取玻璃瓶中的气体3 min,接着通入新鲜空气,使玻璃瓶中的气体与外界的气体平衡,再次进行上述操作,如此反复3次,确保玻璃瓶中充满新鲜空气。在最后一次通入空气时,采集此时的空气于20 mL真空瓶中,作为初始气体浓度,记录采样时间。密封培养6 h之后,从每组处理中随机取出3个土壤样品作为重复,用注射器快速反复抽提5次以确保玻璃瓶内气体充分混匀,用带有三通阀的注射器立即采集20 mL气体,注入20 mL的真空瓶中用于测定N2O和CO2的浓度。
分别在培养过程中的第1、3、7、14、30 d,从每组处理中随机取出3个土壤样品作为重复,按水土比5:1向玻璃瓶中加入100 mL 2 mol·L-1 KCl溶液,振荡,过滤,收集滤液于塑料瓶中,并于4 ℃下低温保存,用于测定土壤中可溶性全氮(DTN)、NO3- -N和NH4+ -N浓度。
1.3 测定项目与方法土壤KCl溶液提取后的滤液用Skalar连续流动分析仪测定DTN、NO3--N和NH4+-N浓度;N2O和CO2的浓度采用气相色谱仪(Agilent Technologies7890A)测定。土壤pH用电位法(KCl浸提液)测定。
1.4 数据分析处理净矿化速率按照公式(1)进行计算:
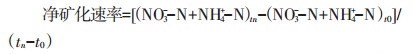
|
(1) |
式中:(NO3--N+NH4+-N)tn和(NO3--N+NH4+-N)t0分别为培养第n d和第0 d时的NO3--N和NH4+-N含量之和,mg·kg-1;tn为培养第30 d;t0为培养第0 d。
净硝化速率按照公式(2)进行计算:

|
(2) |
式中:(NO3--N)t0和(NO3--N)tn分别为培养第n d和第0 d时的NO3--N含量,mg·kg-1;tn为培养第30 d;t0为培养第0 d。

|
(3) |
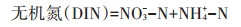
|
(4) |

|
(5) |
式中:F为气体的排放速率(N2O,μg N·kg-1·d-1;CO2,mg C·kg-1·d-1);ρ为标准状况下CO2和N2O的密度(N2O-N,kg·m-3;CO2-C,kg·m-3);dc/dt为单位时间内玻璃瓶内气体浓度增加量,mg·L-1·d-1;V为玻璃瓶中气体的有效空间体积,mL;T为培养温度,℃;W为干土的质量,kg。两次排放通量测定间隔时间内的N2O和CO2排放量用两次测定的平均排放通量乘以时间间隔计算。整个培养过程中的N2O和CO2总的排放量则为N2O和CO2累积排放量。
结果中的可溶性氮组分、N2O和CO2排放数据均为3次重复数据。采用SPSS 24.0软件对数据进行单因素方差分析,用LSD方法(P < 0.05)分析处理间平均数的差异显著性,用OriginPro 2018进行作图和线性拟合。
2 结果与分析 2.1 可溶性氮 2.1.1 铵态氮(NH4+-N)添加黄腐酸钾处理的NH4+-N含量变化趋势与CK处理的NH4+-N含量变化趋势基本一致(图 1A),在前3 d内小幅度减小,随后大幅度降低,第14 d后基本稳定在较低水平。与CK处理相比,添加黄腐酸钾处理在前7 d内显著降低了土壤NH4+-N含量(P < 0.05),降低幅度随黄腐酸钾添加量的增大而增大。

|
CK,硝酸铵;T1,硝酸铵+2.5 g·kg-1黄腐酸钾;T2,硝酸铵+5 g·kg-1黄腐酸钾;T3,硝酸铵+10 g·kg-1黄腐酸钾;T4,硝酸铵+15 g·kg-1黄腐酸钾。误差棒长短表示标准差大小(n=3)。下同 CK, ammonium nitrate; T1, ammonium nitrate +2.5 g·kg-1 potassium fulvic acid; T2, ammonium nitrate +5 g·kg-1 potassium fulvic acid; T3, ammonium nitrate +10 g·kg-1 potassium fulvic acid; T4, ammonium nitrate +15 g·kg-1 potassium fulvic acid. Bars represent standard deviations(n=3). The same below 图 1 添加黄腐酸钾对土壤铵态氮(A)、硝态氮(B)、无机氮(C)和可溶性有机氮(D)含量的影响 Figure 1 Effects of fulvic acid potassium addition on the concentrations of ammonium nitrogen(A), nitrate nitrogen(B), inorganic nitrogen(C)and soluble organic nitrogen(D)in soil |
添加黄腐酸钾处理的NO3--N含量变化趋势与CK处理的NO3--N含量变化趋势不一致(图 1B),CK处理呈不断增大的趋势,而添加黄腐酸钾处理基本上呈先减小后增大到一定值后再次减小的趋势。在培养结束时,T1、T2、T3、T4处理的NO3--N含量显著低于CK处理(P < 0.05),较CK处理分别降低了38.33、49.70、94.73 mg·kg-1和144.95 mg·kg-1。可见,添加黄腐酸钾可显著降低土壤NO3--N含量(P < 0.05),且降低幅度随黄腐酸钾添加量的增大而增大。
图 2B为各处理的净硝化速率,添加黄腐酸钾处理的净硝化速率显著低于CK处理(P < 0.05)。T1、T2、T3、T4处理的净硝化速率分别为2.25、1.87、0.37 mg·kg-1·d-1和-1.31 mg·kg-1·d-1,而CK处理的净硝化速率为3.53 mg·kg-1·d-1。可见,添加黄腐酸钾显著降低了土壤净硝化速率,且降低程度也随黄腐酸钾添加量的增加而增大。

|
不同小写字母表示处理间在0.05水平的显著差异性。下同 Different lowercase letters mean insignificant difference between treatments based on LSD test(P<0.05). The same below 图 2 配施黄腐酸钾对土壤净矿化速率(A)和净硝化速率(B)的影响 Figure 2 Effects of application of fulvic acid potassium on soil net mineralization rate(A)and net nitrification rate(B) |
DIN主要包括NO3--N和NH4+-N。由图 1C和图 1B可以看出,在培养过程中DIN的变化趋势与NO3--N的变化趋势基本一致。整个培养过程中,添加黄腐酸钾处理的DIN含量均小于CK处理,除了T1处理在第14 d比CK处理增大了2.52 mg·kg-1。与CK处理相比,添加黄腐酸钾处理在培养结束时显著降低了土壤DIN含量(P < 0.05),降低幅度以T4处理最大(67.22%),其次为T3处理(44.11%)、T2处理(23.18%)、T1处理(17.78%)。T1、T2、T3、T4处理的净矿化速率分别为-1.20、-1.58、-3.08 mg·kg-1·d-1和-4.73 mg·kg-1·d-1,而CK处理的净矿化速率为0.08 mg·kg-1·d-1(P < 0.05,图 2A)。可以看出,添加黄腐酸钾显著降低了土壤氮素的净矿化速率。
2.1.4 可溶性有机氮(DON)由图 1D可以看出,添加黄腐酸钾可以不同程度地提高土壤DON含量。与CK处理相比,T1和T2处理在前7 d内降低了土壤DON含量,随后又提高了土壤DON含量;T3处理仅在培养开始和结束时显著提高了土壤DON含量;而T4处理在整个培养过程中均显著提高了土壤DON含量。在培养结束时,T1、T2、T3、T4处理分别比CK处理增加了1.24、1.11、3.88 mg·kg-1和7.46 mg·kg-1。可见,当黄腐酸钾的添加量≥10 g·kg-1时,可显著提高DON含量(P < 0.05)。
2.2 N2O和CO2排放量图 3A为各处理土壤呼吸速率的变化趋势。可以看出,添加黄腐酸钾明显提高了土壤呼吸速率。T1和T2处理的呼吸速率在前3 d内达到最大值,分别为66.67 mg C·kg-1·d-1和117.72 mg C·kg-1·d-1;而T3和T4处理的呼吸速率在第1 d就达到最大值,分别为252.16 mg C·kg-1·d-1和400.09 mg C·kg-1·d-1,说明呼吸速率达到最大值的时间随黄腐酸钾添加量的增加而缩短。T1、T2、T3、T4处理的呼吸速率达到最大值后均呈现不断减小的趋势。CK处理的呼吸速率变化范围为11.75~26.43 mg C·kg-1·d-1,在整个培养过程中均显著小于添加黄腐酸钾处理的呼吸速率(P < 0.05)。另外,由图 4A可以看出,T1、T2、T3、T4处理显著提高了土壤CO2累积排放量(P < 0.05),分别比CK处理提高了738.11、1 138.86、1 930.33 mg C·kg-1和2 597.92 mg C·kg-1。

|
图 3 添加黄腐酸钾对CO2(A)和N2O(B)排放速率的影响 Figure 3 Effects of fulvic acid potassium addition on CO2(A)and N2O(B)emission rates |

|
图 4 添加黄腐酸钾对CO2 A)和N2O(B)累积排放量的影响 Figure 4 Effects of fulvic acid potassium addition on cumulative emissions of CO2(A)and N2O(B) |
图 3B为各处理N2O排放速率的变化趋势。T1、T2、T3、T4处理的N2O排放速率在前3 d内达到最大值,分别为2 448.22、3 512.57、7 259.92 µg N·kg-1·d-1和7 393.66 µg N·kg-1·d-1,之后均呈不断减小的趋势。在培养结束时,T1、T2、T3、T4处理的N2O排放速率分别降至1.14、0、0、0 µg N·kg-1·d-1。CK处理的N2O排放速率在整个培养过程中均显著小于添加黄腐酸钾处理(P < 0.05),变化范围为0~27.04 µg N·kg-1·d-1。可见,添加黄腐酸钾显著提高了N2O排放速率(P < 0.05),且N2O排放速率随黄腐酸钾添加量的增加而增大。另外,添加黄腐酸钾处理较CK处理显著增加了N2O累积排放量(P < 0.05,图 4B),且添加黄腐酸钾处理间差异显著。
通过对CO2累积排放量和N2O累积排放量进行线性拟合发现,两者呈显著正相关关系(R2=0.97,P < 0.001),线性关系为y=-0.01x-8.95(图 5A)。另外对净硝化速率与N2O累积排放量进行线性拟合发现,两者呈显著负相关关系(R2=0.92,P < 0.001),线性关系为y=-6.26+22.46(图 5B)。

|
图 5 N2O累积排放量与CO2累积排放量(A)和净硝化速率(B)的相关关系 Figure 5 Relationships between N2O cumulative emissions and CO2 cumulative emissions(A)and net nitrification rate(B) |
土壤中的NH4+-N和NO3--N被微生物同化,成为生物体有机组成部分,称为无机氮的微生物固持,又称微生物同化作用[39]。有研究表明,向土壤中添加外源碳,如葡萄糖、蔗糖、纤维素、木屑和秸秆等碳源,可促进微生物的活性[40-44]。添加外源碳刺激微生物活性的同时也促进了微生物对氮的需求[45],即微生物对有效氮进行同化作用[40-42]。巨晓棠等[24]研究表明,添加高C/N有机物料(小麦秸秆)显著降低了土壤中无机氮含量;而添加低C/N有机物料(苜蓿秸秆或鸡粪)显著提高了土壤中无机氮含量。另有研究表明,当土壤有机碳源不足时,微生物对NH4+-N的吸收同化能力强于对NO3--N的吸收同化能力;当有充足的有机碳源时,微生物对这两种形态氮素的固定能力几乎相当[46]。本文结果显示,在培养第1 d,T1、T2、T3、T4处理的NH4+ - N含量较CK处理分别降低了14.95、34.55、42.14 mg·kg-1和52.14 mg·kg-1(图 1A),这可能是NH4+-N通过硝化作用转换为NO3--N,但T1、T2、T3、T4处理的NO3--N含量并未增加,反而较CK处理也分别降低了7.25、24.75、16.13、18.49 mg·kg-1;另外,降低的NH4+-N也可能是在硝化过程和反硝化过程中以N2O-N形式释放,但T1、T2、T3、T4处理在培养第1 d的N2O累积排放量仅分别为2.5、3.51、7.26、6.49 mg· kg-1,综上,说明添加黄腐酸钾刺激了微生物对NH4+-N的同化。在培养第3 d,T1、T2、T3、T4处理较CK处理显著降低了NH4+-N和NO3--N含量,而N2O-N只能解释NO3--N降低量中的很小比例,说明向土壤中加入黄腐酸钾,相当于以外源方式增加了微生物可利用的碳源,刺激了微生物对NH4+-N和NO3--N的同化。另外,NH4+-N和NO3--N含量的降低值随着黄腐酸钾添加量的增大而增大,说明微生物同化作用随碳源含量的增加而增大,有效碳含量影响微生物同化作用的强弱[29, 40]。在培养3 d后,从CK处理的NH4+-N和NO3--N含量变化趋势可以看出,此时硝化速率较大,说明此阶段内硝化作用占主导,这解释了添加黄腐酸钾处理在培养3 d后NO3--N含量不断增大的原因。在培养结束时,与CK处理相比,添加黄腐酸钾处理显著降低了无机氮含量,这与马红亮等[47]的研究结果中无机氮含量对外源碳的响应一致。添加黄腐酸钾处理的净矿化速率均为负值(图 2A),说明加入黄腐酸钾对微生物固持的刺激作用大于对矿化作用的刺激作用,表现为微生物对氮素的净固持;另外净硝化速率随黄腐酸钾添加量的增大而减小,甚至T4处理的净硝化速率为负值,这说明添加外源碳不仅影响了矿化作用,也抑制了硝化作用,在其他研究中也发现了相似的结果[10-13]。无机氮含量的降低值远大于N2O累积排放量,也充分说明添加黄腐酸钾可以通过刺激微生物同化作用来固持一定量的无机氮。
土壤氮素转化与有效碳源等因子有关[48]。在培养结束时,添加黄腐酸钾显著降低了土壤中的无机氮含量(图 1C),但增加了可溶性有机氮含量(图 1D),这可从外源碳的加入使土壤中微生物活性增大,大量的碳水化合物被微生物快速分解,甚至外源碳添加促进土壤中顽固性老有机碳的分解得到解释[49-50]。
朱霞等[51]研究表明,可溶性碳的添加能间接增强土壤微生物反硝化作用,从而促进土壤N2O排放。Burford等[52]发现反硝化速率与水溶性碳含量呈极显著相关,土壤中水溶性碳含量越多,反硝化速率越快,土壤N2O排放量也大幅增加。简言之,碳含量决定土壤硝化和反硝化量的大小[53]。从本研究结果中也可以看出,当氮肥和黄腐酸钾同时施入土壤后,土壤N2O排放速率显著增加(P < 0.05,图 3B)。在培养初始,T1、T2、T3、T4处理的N2O排放速率分别为2 448.22、3 512.57、7 259.92、6 491.52 µg N·kg-1,分别是CK处理的103、148、306、274倍,说明向土壤中加入黄腐酸钾,相当于以外源方式增加了微生物可利用的有效碳源,这不仅刺激了微生物同化作用,而且刺激了反硝化作用。续勇波等[54]研究也表明,易矿化的有机碳含量越高,反硝化过程中N2O排放速率越大,N2O被进一步还原的速率变小,N2O总排放量增大。本实验得出相同结果,N2O累积排放量随着黄腐酸钾添加量的增大而增大(图 4B),且添加黄腐酸钾处理的N2O累积排放量均显著高于CK处理(P < 0.05)。另外,Gillam等[30]研究发现,N2O和N2排放量增加通常与土壤中NO3--N浓度增加有关,因为NO3--N是反硝化过程中作为N2O生成的重要电子受体。但本实验得出相反的结果,发现净硝化速率与N2O累积排放量呈显著负相关关系(图 5B)。这可能与本实验在开始时向土壤中加入等量的NH4+-N、NO3--N和黄腐酸钾刺激了微生物对NO3--N同化有关。
Tilston等[41]研究表明,添加外源碳可以提高微生物活性,与我们不同的是他们所用碳源是蔗糖和木屑。本实验结果显示,添加黄腐酸钾处理的土壤呼吸速率显著大于CK处理(图 3A),且土壤呼吸速率随黄腐酸钾添加量的增大而增大。另外,添加黄腐酸钾处理较CK处理显著增加了CO2累积排放量(图 4A),且黄腐酸钾添加量与CO2累积排放量呈极显著正相关(r=0.98,sig=0.000),说明添加黄腐酸钾提高了微生物活性,且微生物活性随黄腐酸钾添加量的增大而增强。芦思佳等[55]研究结果中也表明CO2累积排放量随可溶性碳含量的增大而增大。另外,微生物呼吸作用增强,加快了土壤中O2的消耗,容易形成厌氧环境而促进反硝化作用[51]。因此,微生物呼吸作用增强也会导致N2O排放量的增加。从本实验结果也可以看出,N2O累积排放量与CO2累积排放量呈显著正相关关系(P < 0.001,图 5A),这与Huang等[56]研究结果中N2O累积排放量与CO2累积排放量的关系一致。
综上所述,在室内培养试验条件下,配施黄腐酸钾对植烟土壤氮素转化具有一定影响。但在种植烟草的田间条件下,烟株生长会吸收利用一部分氮素,根系分泌物也会对氮素转化产生一定影响[57],氮素还会发生径流、淋洗损失等,黄腐酸钾对氮素转化的影响会与室内培养试验有一定差异。因此,大田种植烟草条件下黄腐酸钾对氮素转化的影响需做进一步深入研究。另外,添加黄腐酸钾可以明显降低土壤中无机氮含量,那么在烤烟生长后期添加黄腐酸钾是否可以作为一种调控植烟土壤后期氮素供应过多的措施?值得我们开展下一步工作去探究。
4 结论(1)黄腐酸钾作为一种高C/N的碳源,对微生物同化作用的刺激大于对矿化作用的刺激;添加黄腐酸钾可显著降低土壤中的无机氮含量,且无机氮含量的降低值随黄腐酸钾添加量的增大而增大。同时,添加黄腐酸钾也刺激了反硝化作用,显著提高了N2O累积排放量,且反硝化作用随黄腐酸钾添加量的增大而增强。
(2)向土壤中添加黄腐酸钾能够刺激微生物活性,显著提高土壤呼吸速率和CO2累积排放量。另外,CO2累积排放量与N2O累积排放量之间存在显著的正相关关系,因此,CO2累积排放量可作为量化N2O累积排放量的辅助指标。
| [1] |
李春俭, 张福锁, 李文卿, 等. 我国烤烟生产中的氮素管理及其与烟叶品质的关系[J]. 植物营养与肥料学报, 2007, 13(2): 331-337. LI Chun-jian, ZHANG Fu-suo, LI Wen-qing, et al. Nitrogen management and its relation to leaf quality in production of flue-cured tobacco in China[J]. Journal of Plant Nutrition and Fertilizer, 2007, 13(2): 331-337. |
| [2] |
Rosa M, Fabio C, Renato C. Nitrogen requirements for flue-cured tobacco[J]. Agronomy Journal, 2006, 98(3): 666-673. |
| [3] |
刘青丽, 张云贵, 焦永鸽, 等. 西南烟区氮素供应与烤烟氮素吸收的关系[J]. 植物营养与肥料学报, 2017, 23(3): 757-764. LIU Qing-li, ZHANG Yun-gui, JIAO Yong-ge, et al. Relationship between nitrogen supply and nitrogen absorption of flue-cured tobacco in southwest China[J]. Journal of Plant Nutrition and Fertilizer, 2017, 23(3): 757-764. |
| [4] |
巨晓棠, 晁逢春, 李春俭, 等. 土壤后期供氮对烤烟产量和烟碱含量的影响[J]. 中国烟草学报, 2003, 9(增刊): 48-53. JU Xiao-tang, CHAO Feng-chun, LI Chun-jian, et al. The yield and nicotine content of flue-cured tobacco as affected by soil nitrogen mineralization in late stage[J]. Journal of China Tobacco, 2003, 9: 48-53. |
| [5] |
刘青丽, 任天志, 李志宏, 等. 我国主要植烟土壤氮素矿化潜力研究[J]. 植物营养与肥料学报, 2009, 15(6): 1266-1272. LIU Qing-li, REN Tian-zhi, LI Zhi-hong, et al. Nitrogen mineralization potential of major soil planting tobacco in China[J]. Journal of Plant Nutrition and Fertilizer, 2009, 15(6): 1266-1272. |
| [6] |
谷海红, 刘宏斌, 王树会, 等. 应用15N示踪研究不同来源氮素在烤烟体内的累积和分配[J]. 中国农业科学, 2008, 41(9): 2693-2702. GU Hai-hong, LIU Hong-bin, WANG Shu-hui, et al. Study on accumulation and distribution of different sources of nitrogen in flue-cured tobacco plant by 15N tracer[J]. Scientia Agricultura Sinica, 2008, 41(9): 2693-2702. |
| [7] |
刘青丽, 任天志, 李志宏, 等. 植烟黄壤供氮特征研究[J]. 中国农业科学, 2010, 43(1): 87-95. LIU Qing-li, REN Tian-zhi, LI Zhi-hong, et al. The characteristics of nitrogen supply in yellow soil planted with tobacco[J]. Scientia Agricultura Sinica, 2010, 43(1): 87-95. |
| [8] |
朱兆良, 文启孝. 中国土壤氮素[M]. 南京: 江苏科学技术出版社, 1992. ZHU Zhao-liang, WEN Qi-xiao. Nitrogen in soils of China[M]. Nanjing: Jiangsu Science and Technology Press, 1992. |
| [9] |
范晓晖, 朱兆良. 旱地土壤中的硝化-反硝化作用[J]. 土壤通报, 2002, 33(5): 385-391. FAN Xiao-hui, ZHU Zhao-liang. Nitrification and denitrification in upland soils[J]. Chinese Journal of Soil Science, 2002, 33(5): 385-391. |
| [10] |
张乐, 何红波, 章建新, 等. 不同用量葡萄糖对土壤氮素转化的影响[J]. 土壤通报, 2008, 39(4): 775-778. ZHANG Le, HE Hong-bo, ZHANG Jian-xin, et al. Effect of glucose addition with different amount on extraneous nitrogen transformation in soil[J]. Chinese Journal of Soil Science, 2008, 39(4): 775-778. |
| [11] |
朱兆良. 农田中氮肥的损失与对策[J]. 生态环境学报, 2000, 9(1): 1-6. ZHU Zhao-liang. Loss of fertilizer N from plants-soil system and the strategies and techniques for its reduction[J]. Ecology and Environmental Sciences, 2000, 9(1): 1-6. |
| [12] |
贾俊仙, 李忠佩, 车玉萍. 添加葡萄糖对不同肥力红壤性水稻土氮素转化的影响[J]. 中国农业科学, 2010, 43(8): 1617-1624. JIA Jun-xian, LI Zhong-pei, CHE Yu-ping. Effects of glucose addition on N transformations in paddy soils with a gradient of organic C content in subtropical China[J]. Scientia Agriculture Sinica, 2010, 43(8): 1617-1624. |
| [13] |
贾俊仙, 李忠佩, 车玉萍. 添加葡萄糖对不同肥力黑土氮素转化的影响[J]. 土壤学报, 2011, 48(1): 207-211. JIA Jun-xian, LI Zhong-pei, CHE Yu-ping. Effect of glucose addition on nitrogen transformation in black soils different in organic carbon content[J]. Acta Pedologica Sinica, 2011, 48(1): 207-211. |
| [14] |
Groffman P M. Carbon additions increase nitrogen availability in northern hardwood forest soils[J]. Biology and Fertility of Soils, 1999, 29(4): 430-433. |
| [15] |
Magill A H, Aber J D. Variation in soil net mineralization rates with dissolved organic carbon additions[J]. Soil Biology and Biochemistry, 2000, 32(5): 597-601. |
| [16] |
Liang L L, Eberwein J R, Allsman L A, et al. Regulation of CO2 and N2O fluxes by coupled carbon and nitrogen availability[J]. Environmental Research Letters, 2015, 10(3): 034008. |
| [17] |
Hadas A, Kautsky L, Goek M, et al. Rates of decomposition of plant residues and available nitrogen in soil, related to residue composition through simulation of carbon and nitrogen turnover[J]. Soil Biology and Biochemistry, 2004, 36(2): 255-266. |
| [18] |
Moritsuka N, Yanai J, Mori K, et al. Biotic and abiotic processes of nitrogen immobilization in the soil-residue interface[J]. Soil Biology and Biochemistry, 2004, 36(7): 1141-1148. |
| [19] |
Qiu S, McComb A J, Bell R W, et al. Measuring microbial uptake of nitrogen in nutrient-amended sandy soils:A mass-balance based approach[J]. Soil Biology and Biochemistry, 2007, 39(2): 581-589. |
| [20] |
Manuel Montano N, Garcia-Oliva F, Jaramillo V J, et al. Dissolved organic carbon affects soil microbial activity and nitrogen dynamics in a Mexican tropical deciduous forest[J]. Plant and Soil, 2007, 295(1/2): 265-277. |
| [21] |
Farrell M, Prendergast-Miller M, Jones D L, et al. Soil microbial organic nitrogen uptake is regulated by carbon availability[J]. Soil Biology and Biochemistry, 2014, 77: 261-267. |
| [22] |
Prober S M, Thiele K R, Lunt I D, et al. Restoring ecological function in temperate grassy woodlands:Manipulating soil nutrients, exotic annuals and native perennial grasses through carbon supplements and spring burns[J]. Journal of Applied Ecology, 2005, 42(6): 1073-1085. |
| [23] |
Szili-Kovacs T, Torok K, Tilston E L, et al. Promoting microbial immobilization of soil nitrogen during restoration of abandoned agricultural fields by organic additions[J]. Biology and Fertility of Soils, 2007, 43(6): 823-828. |
| [24] |
巨晓棠, 刘学军, 张福锁. 尿素与DCD和有机物料配施条件下氮素的转化和去向[J]. 中国农业科学, 2002, 35(2): 181-186. JU Xiao-tang, LIU Xue-jun, ZHANG Fu-suo. Nitrogen transformation and fate in soil under the conditions of mixed application of urea with DCD or different organic materials[J]. Scientia Agriculture Sinica, 2002, 35(2): 181-186. |
| [25] |
Shi W, Miller B E, Star k J M, et al. Microbial nitrogen transformations in response to treated dairy waste in agricultural soils[J]. Soil Science Society of America Journal, 2004, 68(6): 1867-1874. |
| [26] |
Zhang J B, Zhu T B, Meng T Z, et al. Agricultural land use affects nitrate production and conservation in humid subtropical soils in China[J]. Soil Biology and Biochemistry, 2013, 62: 107-114. |
| [27] |
Burger M, Jackson L E. Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems[J]. Soil Biology and Biochemistry, 2003, 35(1): 29-36. |
| [28] |
Trinsoutrot I, Recous S, Mary B, et al. C and N fluxes of decomposing 13C and 15N Brassica napus L.:Effects of residue composition and N content[J]. Soil Biology and Biochemistry, 2000, 32(11): 1717-1730. |
| [29] |
Cheng Y, Wang J, Wang J Y, et al. The quality and quantity of exogenous organic carbon input control microbial NO3-immobilization:A meta-analysis[J]. Soil Biology and Biochemistry, 2017, 115: 357-363. |
| [30] |
Gillam K M, Zebarth B J, Burton D L, et al. Nitrous oxide emissions from denitrification and the partitioning of gaseous losses as affected by nitrate and carbon addition and soil aeration[J]. Can J Soil Sci, 2008, 88(2): 133-143. |
| [31] |
Lal R. Soil carbon sequestration impacts on global climate change and food security[J]. Science, 2004, 304(5677): 1623-1627. |
| [32] |
Morley N, Baggs E M. Carbon and oxygen controls on N2O and N2 production during nitrate reduction[J]. Soil Biology and Biochemistry, 2010, 42(10): 1864-1871. |
| [33] |
强学彩, 袁红莉, 高旺盛. 秸秆还田量对土壤CO2释放和土壤微生物量的影响[J]. 应用生态学报, 2004, 15(3): 469-472. QIANG Xue-cai, YUAN Hong-li, GAO Wang-sheng. Effect of cropresidue incorporation on soil CO2 emission and soil microbial biomass[J]. Chinese Journal of Applied Ecology, 2004, 15(3): 469-472. |
| [34] |
Kemmitt S J, Lanyon C V, Waite I S, et al. Mineralization of native soil organic matter materials is not regulated by the size, activity or composition of the soil microbial biomass:A new perspective[J]. Soil Biology and Biochemistry, 2008, 40: 61-73. |
| [35] |
Shahid M, Dumat C, Silvestre J, et al. Effect of fulvic acids on lead induced oxidative stress to metal sensitive Vicia faba L. plant[J]. Biology and Fertility of Soils, 2012, 48(6): 689-697. |
| [36] |
Priya B N V, Mahavishnan K, Gurumurthy D S, et al. Fulvic acid (FA) for enhanced nutrient uptake and growth:Insights from biochemical and genomic studies[J]. Journal of Crop Improvement, 2014, 28(6): 740-757. |
| [37] |
张亚飞, 罗静静, 彭福田, 等. 黄腐酸钾与化肥控释袋促进桃树生长及氮肥吸收利用[J]. 植物营养与肥料学报, 2017, 23(4): 998-1005. ZHANG Ya-fei, LUO Jing-jing, PENG Fu-tian, et al. Fulvic acid potassium and nutrient-controlled release paperbag enhance peach tree growth, nitrogen absorption and utilization[J]. Plant Nutrition and Fertilizer Science, 2017, 23(4): 998-1005. |
| [38] |
彭玲, 刘晓霞, 何流, 等. 不同黄腐酸用量对'红将军'苹果产量、品质和15N-尿素去向的影响[J]. 应用生态学报, 2018, 29(5): 1412-1420. PENG Ling, LIU Xiao-xia, HE Liu, et al. Effects of different fulvic acid application rates on fruit yield, quality and fate of 15N-urea in 'R ed General' apple[J]. Chinese Journal of Applied Ecology, 2018, 29(5): 1412-1420. |
| [39] |
程谊, 张金波, 蔡祖聪. 土壤中无机氮的微生物同化和非生物固定作用研究进展[J]. 土壤学报, 2012, 49(5): 1030-1036. CHENG Yi, ZHANG Jin-bo, CAI Zu-cong. A research progress on biotic and abiotic inorganic N immobilization in soils[J]. Acta Pedologica Sinica, 2012, 49(5): 1030-1036. |
| [40] |
Sawada K, Funakawa S, Toyota K, et al. Potential nitrogen immobilization as influenced by available carbon in Japanese arable and forest soils[J]. Soil Science and Plant Nutrition, 2015, 61(6): 917-926. |
| [41] |
Tilston E, Szili-Kovács T, Hopkins D W. Contributions of labile and resistant organic materials to the immobilization of inorganic soil N when used in the restoration of abandoned agricultural fields[J]. Soil Use and Management, 2010, 25(2): 168-174. |
| [42] |
Rasul G, Khan A A, Khan K S, et al. Immobilization and mineralization of nitrogen in a saline and alkaline soil during microbial use of sugarcane filter cake amended with glucose[J]. Biology and Fertility of Soils, 2009, 45(3): 289-296. |
| [43] |
Blagodatskaya E, Khomyakov N, Myachina O, et al. Microbial interactions affect sources of priming induced by cellulose[J]. Soil Biology and Biochemistry, 2014, 74: 39-49. |
| [44] |
Kelliher F M, Barbour M M, Hunt J E. Sucrose application, soil microbial respiration and evolved carbon dioxide isotope enrichment under contrasting land uses[J]. Plant and Soil, 2005, 268(1): 233-242. |
| [45] |
Chen R, Senbayram M, Blagodatsky S, et al. Soil C and N availability determine the priming effect:Microbial N mining and stoichiometric decomposition theories[J]. Global Change Biology, 2014, 20(7): 2356-2367. |
| [46] |
Szili-Kovács T, Török K, Tilston E L, et al. Promoting microbial immobilization of soil nitrogen during restoration of abandoned agricultural fields by organic additions[J]. Biology and Fertility of Soils, 2007, 43(6): 823-828. |
| [47] |
马红亮, 魏春兰, 李磊, 等. 添加葡萄糖对杉木人工林土壤氮素转化及净矿化和硝化的影响[J]. 林业科学研究, 2014, 27(3): 356-362. MA Hong-liang, WEI Chun-lan, LI Lei, et al. Effects of glucose addition on soil nitrogen transformation and net mineralization and nitrification in Chinese fir plantation[J]. Forest Research, 2014, 27(3): 356-362. |
| [48] |
方运霆, 莫江明, Per Gundersen, 等. 森林土壤氮素转换及其对氮沉降的响应[J]. 生态学报, 2003, 24(7): 211-219. FANG Yun-ting, MO Jiang-ming, Per Gundersen, et al. Nitrogen transformations in forest soils and its responses to atmospheric nitrogen deposition:A review[J]. Acta Ecologica Sinica, 2003, 24(7): 211-219. |
| [49] |
Fontaine S, Barot S, Barré P, et al. Stability of organic carbon in deep soil layers controlled by fresh carbon supply[J]. Nature, 2007, 450(7167): 277-280. |
| [50] |
Miao S J, Ye R Z, Qiao Y F, et al. The solubility of carbon inputs affects the priming of soil organic matter[J]. Plant and Soil, 2017, 410(1/2): 129-138. |
| [51] |
朱霞, 韩晓增, 乔云发, 等. 外加可溶性碳氮对不同热量带土壤N2O排放的影响[J]. 农业环境科学学报, 2009, 28(12): 2637-2644. ZHU Xia, HAN Xiao-zeng, QIAO Yun-fa, et al. Influence of soluble carbon and nitrogen on N2O emission from different thermal zones soil[J]. Journal of Agro-Environment Science, 2009, 28(12): 2637-2644. |
| [52] |
Burford J R, Bremner J M. Relationships between the denitrification capacities of soils and total, water-soluble and readily decomposable soil organic matter[J]. Soil Biology and Biochemisty, 1975, 7(6): 389-394. |
| [53] |
Katz R, Hagin J, Kurtz L T. Participation of soluble and oxidizable soil organic compounds in denitrification[J]. Biology and Fertility of Soils, 1985, 1(4): 209-213. |
| [54] |
续勇波, 蔡祖聪. 亚热带土壤氮素反硝化过程中N2O的排放和还原[J]. 环境科学学报, 2008, 28(4): 731-737. XU Yong-bo, CAI Zu-cong. Nitrous oxide emission and reduction during denitrification in subtropical soils[J]. Acta Scientiae Circumstantiae, 2008, 28(4): 731-737. |
| [55] |
芦思佳, 韩晓增. 施肥对土壤呼吸的影响[J]. 农业系统科学与综合研究, 2011, 27(3): 366-370. LU Si-jia, HAN Xiao-zeng. The effect of fertilization on soil respiration[J]. System Sciences and Comprehensive Studies in Agriculture, 2011, 27(3): 366-370. |
| [56] |
Huang Y, Zou J, Zheng X, et al. Nitrous oxide emissions as influenced by amendment of plant residues with different C:N ratios[J]. Soil Biology & Biochemistry, 2004, 36(6): 973-981. |
| [57] |
俞慎, 李振高. 稻田生态系统生物硝化-反硝化作用与氮素损失[J]. 应用生态学报, 1999, 10(5): 630-634. YU Shen, LI Zhen-gao. Biological nitrification-denitrification and nitrogen loss in rice field ecosystem[J]. Chinese Journal of Applied Ecology, 1999, 10(5): 630-634. |