 2020, Vol. 39
2020, Vol. 39



2. 中国科学院南京地理与湖泊研究所 湖泊与环境国家重点实验室 太湖湖泊生态系统研究站, 南京 210008;
3. 中国科学院大学, 北京 100049
2. Taihu Laboratory for Lake Ecosystem Research, State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
中国是全球最大的渔业养殖和水产品消费国家,水产养殖产量占世界水产养殖总产量的60%以上[1]。而内陆淡水池塘养殖又是其中最主要的养殖方式,2018年养殖面积达252.7万hm2,养殖产量2 212.2万t,占全国水产品总产量的32.9%[2]。池塘养殖时,强调“养鱼先养水”,即鱼苗在下塘养殖前,要提前对养殖池塘的水体进行施肥等管理工作,使得养殖水体保持“肥、活、嫩、爽”。从生态学角度讲,这个过程就是通过施肥加速养殖水体富营养化,促进藻类及浮游动物的生长,从而提高水体生产力,为鱼类提供充足的生物饵料,形成相对稳定并适合鱼类生活的微生态环境。
与自然水域生态系统不同,淡水池塘养殖是特殊的人为操纵生态系统。充足的营养盐通过上行效应提高鱼类产量的同时,大量投入的有机质也为池塘生态系统食物网的腐屑链提供了能量和物质基础。细菌在腐屑链中占据着重要地位,一方面细菌起着降解作用,是物质和能量的贮存者,另一方面,它们是重要的生产者,能够将有机质转化为细菌菌体蛋白,直接被浮游动物和鱼类摄食[3]。已有研究表明,细菌群落与水体的营养水平及浮游植物的生物量存在明显关系[4-5]。养殖池塘中,由于富营养化和鱼类的牧食压力,小型浮游植物和浮游动物占据优势[6],进而可能对池塘生态系统中的细菌群落结构造成影响。因此,对养殖池塘水体细菌群落结构开展研究十分必要。本文采用16S rRNA基因片段高通量测序技术对南京斑点叉尾鮰(Ictalurus punctatus)淡水养殖池塘水体细菌群落的周年变化开展了研究,目的主要是:(1)了解斑点叉尾鮰养殖池塘水体细菌群落组成、多样性的年度变化特点;(2)明确斑点叉尾鮰养殖池塘水体细菌群落结构的主要影响因子。这些研究结果将对斑点叉尾鮰养殖池塘水体的精细化管理和健康养殖提供帮助。
1 材料与方法 1.1 养殖池塘及样品采集试验在江苏省淡水水产研究所国家斑点叉尾鮰遗传育种中心养殖基地开展。养殖池塘面积0.21 hm2(50 m×42 m),位于南京市江宁区禄口镇(31°77′ 03″N,118°88′29″E)。养殖开始前一个月抽干池水,漂白粉清塘后,加水至1.5 m左右,池塘中央设置一台增氧机。4—10月,每晚23:00开启增氧机至次日凌晨4:00,晴天中午11:00点开启2 h,保证养殖水体溶氧充足。夏季每月月中补水,春秋季则每2月补水一次。池塘内放入2 747尾斑点叉尾鮰鱼种(平均18.91 g·尾-1),养殖从11月底至次年12月初。每日早晚两次投喂漂浮性颗粒配合饲料(蛋白含量35%),饲料投喂量为鱼体质量的3%左右。
养殖周期内,每月的月末开展水样采集。每次上午10:00,利用Schindler采水器(5 L)采集3处池塘表层50 cm的水样,放入灭菌塑料瓶。水样放入4 ℃保温箱带回实验室,分析时样品为3个点水样的混合样。取300 mL混合水样经0.22 μm的聚碳酸酯膜(Millipore,Cork,Ireland)负压过滤,得到滤膜样品,放入2 mL的无菌离心管,-80 ℃超低温冰箱冷冻保存,留待DNA提取。另取200 mL混合水样4 ℃低温保存,进行水质的化学因子分析。
1.2 池塘水质因子测定利用多功能水质参数仪(YSI6920v2,USA)现场测定温度(Temp)、酸碱度(pH)、总溶解性悬浮物(TDS)、浊度(Tur)以及溶解氧(DO)等环境因子。塞氏盘测定水体透明度(SD)。其他水质指标在实验室内按以下方法测定:总氮(TN)采用碱性过硫酸钾消解紫外分光光度法(GB 11894—1989);铵态氮(NH4+-N)采用纳氏试剂分光光度法(GB 7479—1987);硝态氮(NO3--N)采用酚二磺酸分光光度法(GB 7480—1987);亚硝态氮(NO2--N)采用分光光度法(GB 7493—1987);总磷(TP)和正磷酸盐(PO43--P)采用磷钼蓝比色法(HJ 670-2013);总有机碳(TOC)采用燃烧-非分散红外吸收法(GB 13193—1991);化学需氧量(CODMn)采用高锰酸钾滴定法(GB 11892—1989)。水样经Whatman GF/C 0.45 μm滤膜过滤后,分光光度法(HJ 897—2017)测定叶绿素a (Chl a)含量。
1.3 DNA提取和16S rRNA基因扩增水样过滤收集滤膜,用预先经75%酒精擦拭过的医用剪刀剪碎后,利用Qiagen DNeasy Blood & Tissue Kit试剂盒(Qiagen,CA,USA)提取微生物总DNA。提取的DNA经紫外分光光度计和1%的琼脂糖凝胶电泳检测浓度及纯度后,-20 ℃保存备用。以通用引物338F和806R扩增16S rRNA基因V3–V4高变区[7],建库及高通量测序在上海美吉生物医药科技有限公司完成。
1.4 数据优化与分析生物信息学分析前,先用Trimmomatic软件[8]去除测序原始序列的引物和barcode标签,再用FLASH软件[9]对序列进行拼接,使用VSEARCH(1.9.6)将序列以97%相似性聚类成操作分类单元(Operational taxonomic units,OTUs),比对的参考数据库是Silva_132 16SrRNA database (http://www.arb-sliva.de) [10]。
所有统计分析及可视化分析均利用R语言中的vegan和ggplot2程序包(Version 3.2.2)以及上海美吉生物医药科技有限公司I-sanger云平台完成。多样性分析前,根据每个样本中最少的序列数进行抽平。利用mothur软件计算α多样性[11]。计算β多样性前,对抽平后的物种数据进行Hellinger转化[12],再进行非度量多维尺度分析(Nonmetric multi-dimensional scaling,NMDS) [13]。对环境因子进行999次Monte Carlo置换检验的前向选择后,采用冗余分析(Redundancy analysis,RDA)对环境因子与微生物群落进行典范对应分析[14]。采用相似性分析(Analysis of similarities,ANOSIM)检验季节间差异。最后,通过多物种线性差异判别分析(Linear discriminant analysis effect size,LEfSe),根据分类学组成对样本按照不同分组条件进行线性判别分析(LDA),找出具有季节显著性差异的菌群[15]。
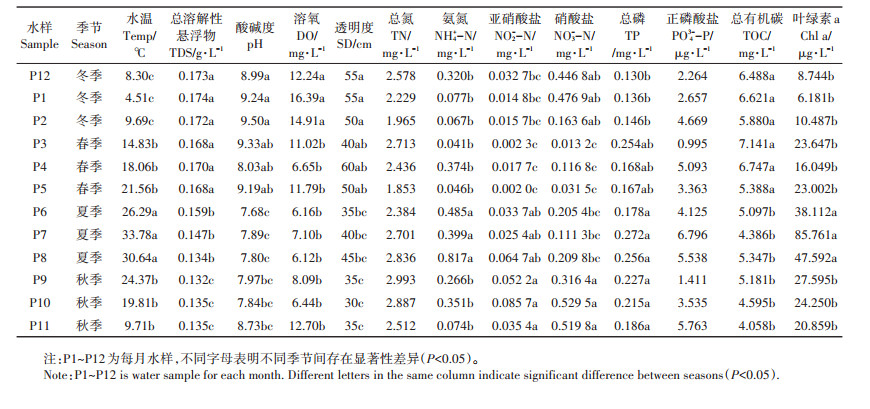
2 结果与分析 2.1 水体理化特征年度养殖试验过程中,水温(Temperature)呈现出明显的季节性变化,从冬季的1月末逐步升高,夏季7月末达到峰值33.78 ℃,之后缓慢下降,除春季和秋季外,其他季节间水温都存在显著性差异(P < 0.05)。总溶解性悬浮物(TDS)和水体透明度(SD)都呈现出持续下降的趋势,且都是冬春季显著高于夏秋季(P < .05)。全年水样pH介于7.68~9.50,始终保持偏碱性,冬季水体的碱性显著强于夏季(P < 0.05)。水体溶氧(DO)含量为6.12~16.39 mg·L-1,适合鱼类养殖,冬季水体的溶氧含量也显著高于其他3个季节(P < .05)。水体总氮(TN)和总磷(TP)浓度始终维持较高水平,一直处于富营养化状态,四季间TN变化不大,但冬季TP浓度显著低于夏秋季节(P < 0.05)。氨氮(NH4+-N)、亚硝酸盐(NO2--N)、硝酸盐(NO3--N)随季节的变化表现出一定的差异,但正磷酸盐(PO43--P)则全年无显著性变化。总有机碳(TOC)也表现为冬春季显著高于夏秋季(P < 0.05)。叶绿素a(Chl a)夏秋季要高于冬春季,其中夏季显著高于其他3个季节,并在7月达到年度峰值,冬季的叶绿素a浓度最低。水体理化特征见表 1。
|
|
表 1 斑点叉尾鮰养殖池塘水体理化性质 Table 1 Summary of physicochemical parameters for water samples of channel catfish pond |
经优化处理后,共获得356 877条序列,平均每个样品有29 741条高质量序列,序列平均长度为434.55 bp。这些序列被聚类为1 023个OTUs。稀释曲线分析中所有样本的都接近平缓,表明本次试验抽样充分,测序深度足以反映每个样本细菌群落的生物信息。
选取Ace指数、Chao1指数、Shannon指数和Simpson指数用来反映养殖池塘水体细菌群落的物种丰富度和多样性。结果显示(图 1),养殖年度内,Ace指数和Chao1指数变化趋势较为一致:冬春季呈S形变化,最小值出现在冬季2月,并与相邻的1月以及初春的3月形成细菌群落物种丰富度的小低谷,之后的3个月又持续上升;7月Ace指数和Chao1指数大幅下降,之后锯齿状震荡上升,并在10月达到年度最大值。使用单因素方差分析(ANOVA)对4个季度的Ace指数和Chao1指数进行多重比较,结果表明,两个指数只有秋季和冬季之间存在显著性差异(P < 0.05)。年度内养殖池塘水体细菌群落的Shannon指数和Simpson指数变化趋势呈相反的趋势,表明细菌群落多样性变化相对稳定,预测结果较为一致。Shannon指数和Simpson指数为对应的“山”字状,7月是细菌群落多样性的最低值,且与其他月份差别较大,3月则是倒数第二低值,其他月份变化都不大(图 1)。单因素方差分析与多重比较(ANOVA)结果显示,4个季度Shannon指数和Simpson指数间无显著性差异(P>0.05)。

|
图 1 斑点叉尾鮰养殖池塘水体微生物群落α多样性 Figure 1 The alpha diversity of bacterial community in channel catfish pond |
使用非度量多维尺度分析(NMDS)展示养殖池塘水样细菌群落的β多样性。NMDS分布图(图 2A)显示,所有样本呈现出较好的季节性排列趋势,即4个季度的样本可以被明显地区分,同一季度内的样品间距离更近,其微生物群落也更加相似。同时,整个年度水体细菌群落也呈现出连续的演替特征,从排序图的左侧连续往右排列,只有夏季样品的群落稍有例外,与秋季略有重叠,但整体排在右侧。NMDS分析的Stress值为0.042,也表明排序结果可靠。细菌群落层次聚类分析的结果与NMDS类似,所有群落聚为两簇(图 2B):夏季和秋季样品(11月样品除外)聚为一簇,冬季和春季的样品聚为另一簇。各分簇内部,每个季节的样品也聚类为一支。

|
图 2 斑点叉尾鮰养殖池塘水体微生物群落NMDS排序图和层级聚类分析 Figure 2 NMDS plot and hierarchical clustering analysis of the compositional dissimilarities of the bacterial communities |
1 023个OUT被注释为27个门(Phyla)、60个纲(Classes)和347个属(Genera)。为了减少群落的复杂性和冗余性,相对丰度低于1%的群落都归为稀有种,并为“Others” [16]。
养殖池塘水体样本中OTU数量从高到低呈现为秋季>春季>夏季>冬季。4个季节样本共享的OUT有324个,占全部OTU的31.67%,3个季节和2个季节样品间共享OUT分别为274个和267个,占比26.78%和26.09%。而单季样品独有的OUT数量最少,只有158个,占比15.44%。其中,冬季和秋季样品独有OUT最多,分别为48个和47个,占比4.69%和4.59%,夏季样品独有OUT最少,仅29个,占比2.83%。
ANOSIM检验表明4个季度养殖池塘水体样品间细菌群落组成存在显著性差异(P < 0.01)。然而,分析细菌群落结构组成,所有样品在门水平上组成较为相似。相对丰度从高到低依次为:蓝藻门Cyanobacteria (29%)、放线菌门Actinobacteria(25%)、变形菌门Proteobacteria (20%)和拟杆菌门Bacteroidetes (12%),这4个门占全部群落的86%。而疣微菌门Verrucomicrobia(3.9%)、浮霉菌门Planctomycetes (3.2%)、厚壁菌门Firmicutes(2.5%)、绿菌门Chlorobi (1.8%)和绿弯菌门Chloroflexi(1.4%)相对丰度都低于4%。
门水平上,冬季水体样品中放线菌门相对丰度最高,之后依次为变形菌门、蓝藻门以及拟杆菌门。而春、夏、秋3季的水体样品中蓝藻门相对丰度最高,之后分别为放线菌门、变形菌门和拟杆菌门。各个季度样品内其他菌门的相对丰度都低于5%。从时间角度上看,养殖池塘水体中4个主要菌门的相对丰度呈现出截然不同的变化趋势(图 3)。蓝藻门的相对丰度呈单峰形变化,夏季达到最高峰(37%),春秋季略有下降(30%和32%),冬季最低(17.45%)。其中,3月和7月水体中蓝藻门相对丰度出现了两次明显的升高,分别为49.97%和56.77%。放线菌门相对含量则表现为增长的趋势,从春季的20%一直增加至冬季的29%。变形菌门1年内的相对丰度保持相对稳定,维持在20%左右。拟杆菌门则呈U字形变化,即冬春季高(15%左右),夏秋季低(8.5%)。

|
图 3 斑点叉尾鮰养殖池塘水体微生门级水平细菌群落结构的年度变动 Figure 3 Annual dynamic of bacterial community composition at the phylum level |
4个季度样品间的细菌群落结构比较显示,厚壁菌门、绿菌门和Saccharibacteria门存在显著性差异(P < .05)。其中,冬春季的厚壁菌门相对数量显著高于夏秋季,绿菌门相对数量则是夏秋季显著高于冬春季,而秋季Saccharibacteria门的相对数量显著高于其他3个季节。
属水平上,养殖池塘水体的优势菌属为hgcI_clade、蓝细菌属norank_c__Cyanobacteria、聚球藻属Synechococcus、CL500-29、CL500-3和多核杆菌属Polynucleobacterium。聚类热图表明,属水平上,所有样品聚为冬春和夏秋两簇(图 4)。其中,肠球菌属Enterococcus、芽孢杆菌属Bacillus、乳球菌属Lactococcus、单胞菌属Polaromonas、红育菌属Rhodoferax和黄杆菌属Flavobacterium的相对丰度在冬春季明显高于夏秋季。而甲基嗜酸菌属Candidatus_Methylacidiphilum、球衣菌属Sphaerotilus、玫瑰单胞菌属Roseomonas、不动杆菌属Acidibacter和湖沉积杆菌属Limnobacter的相对丰度则是夏秋季明显高于冬春季。

|
图 4 养殖池塘水体微生物属级水平的分层聚类热图 Figure 4 Bacterial community heatmap analysis in genus level |
4个季度样品间的细菌群落结构比较在3个菌门发现了显著性差异,在时间维度上进一步采用LefSe软件对养殖水体细菌群落样本进行线性判别分析(LDA),以找到各个季节显著性对应的群落或物种。对LDA判别值大于3.5的细菌类群进行分析,共有不同分类水平的53个细菌类群,在4个季节间具有显著差异(图 5)。春、夏和秋3个季节的显著优势种相对集中,主要分布于一两个门。其中,春季的水体中,优势性菌群主要集中于厚壁菌门中的肠球菌属Enterococcus,以及蓝藻门中的SubsectionIV。夏季水样中,优势性菌群集中于变形菌门中的δ-变形菌纲Deltaproteobacteria,寡弯菌属Oligoflexaceae,以及变形菌门中的球衣菌属Sphaerotilus和蓝藻门中的颤藻属Oscillatoria。秋季的样品优势菌群则主要集中于绿菌门中的OPB56属、Saccharibacteria门,以及蓝藻门中的属FamilyI和变形菌门中的红螺菌目Rhodospirillales与玫瑰单胞菌属Roseomonas。冬季具有显著优势菌群最多,共有2个纲、6个目、9个科和10个属。其中,疣微菌门中的疣微菌纲Verrucomicrobiae疣微菌科Verrucomicrobiaceae、丰佑菌纲Opitutae Vadin中的HA64属、放线菌门中的PeM15纲、从棒状杆菌纲Corynebacteriales中的分枝杆菌属Mycobaterium、厚壁菌门中的芽孢杆菌目Bacillales芽孢杆菌属Bacillus、链球菌科Streptococcaceae乳球菌属Lactococcus、变形菌门中的红细菌目Rhodobacterales到红杆菌属Rhodobacteraceae、MNG科、以及其他零星分布的普雷沃氏菌科Prevotellaceae、球形红杆菌属Candidatus_Aquirestis、嗜甲基菌属Methylotenera、单胞菌属Polaromona和Limnohabitans属。

|
图 5 斑点叉尾鮰养殖池塘四季间差异性菌群多物种线性差异判别分析 Figure 5 LEfSe identified the most differentially abundant taxa in the channel catfish pond among four seasons |
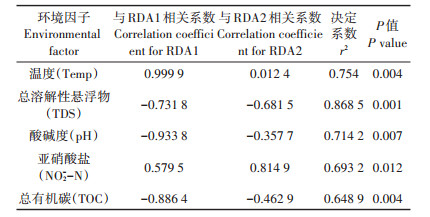
与样本菌群组成相关的环境因子很多,但其中部分因子存在共线性,所以先进行筛选,保留相互作用较小的环境因子。方差膨胀因子(Variance inflation factor,VIF)分析表明自变量间的多重共线性关系严重,过滤掉VIF大于20的环境因子NO3--N和Chl a,多次筛选后,剩下的环境因子对应的VIF值全部小于20。Monte Carlo排序检验表明Temp、TDS、pH、NO2-N、TOC与细菌群落之间存在显著的相关性(P < 0.05)。这5个因子对细菌群落格局的影响大小(r2)依次顺序是:TDS>Temp>pH>NO2--N>TOC (表 2)。
|
|
表 2 环境因子对排序结果的相关性系数及显著性检验值 Table 2 Coefficient correlations between physicochemical parameters and RDA axis |
利用前置检验挑选出来的5个与细菌群落显著相关的环境因子进行RDA排序分析,结果表明,RDA1轴和RDA2轴分别解释了细菌群落变异的44.53%和10.98%,主要菌群被聚为2簇(图 6)。第1簇由冬季和春季样品组成,第2簇由夏季和秋季样品组成,两簇被RDA2轴分隔。其中,第1簇冬春季样本分布比较紧密,主要受pH、TOC和TDS的影响;第2簇夏秋季样本分布较为分散,且夏秋季样品被RDA1轴分隔,主要受NO2--N和Temp的影响。RDA结果显示,斑点叉尾鮰养殖池塘水体微生物具有比较明显的季节性,4个季节的样品沿着RDA1轴从左往右排列。

|
图 6 细菌群落与环境因子之间的RDA分析 Figure 6 RDA bioplot of the distribution of bacterial community and environmental variables |
本文通过分析斑点叉尾鮰养殖池塘水体细菌群落的年度变化过程,阐明斑点叉尾鮰养殖池塘水体细菌群落的变化规律。研究表明,养殖池塘的微生物群落与营养盐、水质、养殖对象及日常管理都具有重要的关系[3]。本研究结果表明,斑点叉尾鮰养殖池塘水体优势菌群为蓝藻门、放线菌门、变形菌门和拟杆菌门,与三疣梭子蟹和吉富罗非鱼养殖塘水体较为相似,但是主要优势菌群的相对丰度排序略有差别[17-18]。不同养殖品种,甚至同一养殖品种的不同养殖阶段,其养殖水体的微生物群落都有明显的差别,这与养殖对象的栖息方式、食性特征以及养殖周期等差异有关[18]。斑点叉尾鮰和吉富罗非鱼养殖过程中投饵量大,产生大量的残饵和粪便,往往造成过量的营养物输入至养殖池塘,同时,斑点叉尾鮰和罗非鱼都是底栖性鱼类,因此优势细菌群落较为相似。
ANOSIM检验表明斑点叉尾鮰养殖池塘水体4个季度样品间细菌群落组成存在显著性差异(P < 0.01),NMDS分析(图 2A)也验证了所有样本呈现出较好的季节性排列趋势。斑点叉尾鮰养殖池塘水体优势菌中,拟杆菌门相对丰度冬春季显著高于夏秋季节,LefSe分析进一步表明,主要差异菌群为冬季的普雷沃氏菌科Prevotellaceae、球形红杆菌属Candidatus_Aquirestis。普雷沃氏菌科Prevotellaceae能够利用和降解蛋白质及各类有机物[19],而斑点叉尾鮰养殖池塘的总有机碳显著高于夏秋季节,为普雷沃氏菌科细菌的繁殖提供了适合的环境。球形红杆菌属Candidatus_Aquirestis广泛分布于内陆高硬度水体,冬春季也会在自然水体中形成明显的生物量高峰,秋冬季则维持较低的丰度[20],与斑点叉尾鮰养殖池塘水体的明显季节性变化一致。
蓝藻门作为广泛分布于各类水体的细菌门类,其丰度状况与水体富营养化程度密切相关。水产养殖环境中,因为营养盐和有机物的密集投入,水体长期维持富营养化状态,蓝藻水华爆发已经变得越来越频繁[21]。斑点叉尾鮰池塘的蓝藻含量虽然没有发现显著的季节性差异,但是其相对丰度依然存在季节性的变化规律。春季3月和夏季7月斑点叉尾鮰养殖池塘出现了两次蓝藻生物量的高峰,蓝藻生物量占总细菌生物量的49.98%和56.77%,同时,3月和7月蓝藻生物量的上升导致池塘水体细菌群落α多样性显著降低。LefSe分析表明,春、夏和秋季蓝藻门的显著优势菌群各不相同。春季优势蓝藻主要是念珠藻目的g__norank_f__FamilyI_o__SubsectionIV属,大都呈丝状,形成厚壁孢子,在长换水周期、高pH、高温和高光照强度等条件下,更具有优势[22]。斑点叉尾鮰养殖池塘水体没有剧烈波动,冬春季pH一直较高,3月末温度已经接近15 ℃,营养盐也一直维持较高浓度,因此在3月末,蓝藻迅速生长,并导致了叶绿素a浓度的上升。之后4月蓝藻大量死亡并氧化分解,消耗水体溶氧并产生乳酸等酸性物质,导致4月池塘溶氧含量降低,pH也略有下降。夏季优势的颤藻属Oscillatoria是常见的淡水蓝藻群体,呈丝状不分枝,会节律性颤动。斑点叉尾鮰养殖池塘水体夏季颤藻属快速生长,成为优势类群,并在7月形成蓝藻水华的最高峰,此时池塘水体叶绿素a浓度也是全年最高,这个过程与开放湖库的夏季蓝藻水华演替较为相似[23]。秋季g__norank_f__FamilyI_o__SubsectionI属的蓝藻替代颤藻属成为优势蓝藻类群,该属归属于蓝藻纲Subsection I,皆为单细胞蓝藻,有空泡,并合成藻胆蛋白,是湖泊、水库等天然水体中秋季常见的水华蓝藻种类[24]。
Monte Carlo检验和RDA排序分析表明斑点叉尾鮰养殖池塘水体细菌群落与环境因子间具有显著的相关性,主要的环境因子包括TDS、Temp、pH、NO2--N和TOC。5个环境因子都表现出明显的季节性差异,冬春季的TDS、pH和TOC显著高于夏秋季,Temp和NO2--N则是夏秋季显著高于冬春季。其中,TDS和Temp对细菌群落的影响最大。TDS能干扰水体的氧传递和降低细菌活性,从而影响硝酸盐、有机物和COD的去除[25]。大量的研究都表明温度对细菌群落组成有重要影响。温度会影响细菌降解有机物、再生营养以及参与微食物链等方面的生物学功能[26-27],同时还对水体浮游植物的演替产生影响,改变水体微生态系统[28],从而驱动细菌群落组成的改变。水温季节性的波动造成栖息环境的季节性变化,进一步推动水体细菌群落组成呈现出季节性的响应[29]。此外,不同菌群对温度的敏感差异,也会导致不同温度下细菌群落结构的差异[30]。斑点叉尾鮰养殖池塘春季和夏季的水温升高提高了蓝藻门的生长速率,并给蓝藻门带来了竞争优势[31],最终与其他因素共同导致斑点叉尾鮰养殖池塘3月和7月的两次蓝藻爆发。
pH被认为是生物化学地球转换过程中重要的调控因子,它能调节水体中离子和微量金属的利用率,从而对细菌的生长起到抑制或者促进的作用[32]。养殖水体光合作用的强弱对pH影响较大,因此pH会随着光照强度和初级生产力的改变呈现出明显的季节性变化。本研究中pH冬春明显高于夏秋,这可能与本试验池塘水体初级生产力相对较低有关,叶绿素a浓度明显低于草鱼、乌鳢和罗非鱼等品种的养殖池塘[33-34]。同时,夏季池塘补水量大,可能也会引起pH的降低。N和P一直被认为是细菌和浮游植物生长的限制性因子[35],TOC也被认为能够直接影响细菌的生物量和组成,从而改变细菌群落结构[36]。然而,斑点叉尾鮰养殖水体一直呈人为的富营养化状态,大部分的营养盐(TN、NH4+-N、NO3--N、TP和PO43--P)都没有对细菌群落结构的塑造起到显著性作用。而NO2--N是水体中氮素存在的一个中间态,本研究池塘溶氧持续充足,整个养殖年度内NO2--N浓度一直维持在较低的状态。因此,NO2--N的低浓度加上夏秋高冬春低的波动,会对细菌群落如亚硝酸盐硝化细菌等产生显著的影响。斑点叉尾鮰养殖池塘水体的C/N [可由TOC/ (NH4+-N+NO2--N+NO3--N)粗略计算]只有7~14,远低于细菌生长的最适范围20~30[37],因此C (主要是TOC)是斑点叉尾鮰养殖池塘细菌生长的限制因子。拟杆菌门的菌群能够高效利用低碳物质[38],冬春季的TOC浓度显著高于夏秋季节(P < 0.05,表 1),因此冬春季的拟杆菌门Bacteroidetes丰度也显著高于夏秋季。
4 结论本研究运用16S rDNA扩增技术研究斑点叉尾鮰养殖池塘微生物群落,结果表明斑点叉尾鮰养殖池塘水体细菌群落结构呈现出季节性变化,优势菌群为蓝藻门、放线菌门、变形菌门和拟杆菌门。环境因子中,总溶解性悬浮物、水温、酸碱度、亚硝酸盐和总有机碳对水体细菌群落结构有显著的影响。但是本研究局限于一个地方的斑点叉尾鮰池塘主养模式,而斑点叉尾鮰养殖已经遍布全国,因此,后续研究仍需进一步针对斑点叉尾鮰的不同养殖模式在不同地域开展大尺度的养殖池塘微生物群落结构比较,从而为不同地域不同模式的斑点叉尾鮰养殖管理提供精确理论指导。
| [1] |
FAO. The state of world fisheries and aquaculture 2018: Meeting the sustainable development goals[R]. Rome: FAO, 2018.
|
| [2] |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 2019中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2019. Bureau of Fisheries MOA, National Fisheries Technology Extension Center, China Society of Fisheries. China fishery statistical yearbook 2019[M]. Beijing: China Agriculture Press, 2019. |
| [3] |
Moriarty D J. The role of microorganisms in aquaculture ponds[J]. Aquaculture, 1997, 151(1): 333-349. |
| [4] |
唐永涛, 赵良杰, 杨洋, 等. 基于16S rDNA比较研究混养三角帆蚌和鲢鳙对池塘养殖水体微生物群落结构的影响[J]. 淡水渔业, 2015(3): 70-77. TANG Yong-tao, ZHAO Liang-jie, YANG Yang, et al. Effects of Hyriopsis cumingii, Hypophthalmichthys molitrix and Aristichthys nobilis polycultured on microbial community structure in aquaculture ponds/enclosures:A comparative study based on 16S rDNA sequences[J]. Freshwater Fisheries, 2015(3): 70-77. DOI:10.3969/j.issn.1000-6907.2015.03.012 |
| [5] |
De Wever A, Muylaert K, Van d G K, et al. Bacterial community composition in Lake Tanganyika:Vertical and horizontal heterogeneity[J]. Applied and Environmental Microbiology, 2005, 71(9): 5029-5037. DOI:10.1128/AEM.71.9.5029-5037.2005 |
| [6] |
朱学宝, 施正峰.中国鱼池生态学研究[M].上海: 上海科学技术出版社, 1995. ZHU Xue-bao, SHI Zheng-feng. Study on the ecology of aquaculture pond in China[M]. Shanghai: Shanghai Scientific & Technical Publishers, 1995. |
| [7] |
Masoud W, Takamiya M, Vogensen F K, et al. Characterization of bacterial populations in Danish raw milk cheeses made with different starter cultures by denaturating gradient gel electrophoresis and pyrosequencing[J]. International Dairy Journal, 2011, 21(3): 142-148. DOI:10.1016/j.idairyj.2010.10.007 |
| [8] |
Bolger A M, Marc L, Bjoern U. Trimmomatic:A flexible trimmer for Illumina sequence data[J]. Bioinformatics, 2014, 30(15): 2114-2120. |
| [9] |
Reyon D, Tsai S Q, Khayter C, et al. FLASH assembly of TALENs for high-throughput genome editing[J]. Nature Biotechnology, 2012, 30(5): 460-465. DOI:10.1038/nbt.2170 |
| [10] |
Quast C, Pruesse E, Yilmaz P, et al. The SILVA ribosomal RNA gene database project:Improved data processing and web-based tools[J]. Nucleic Acids Research, 2013, 41: D590-D596. |
| [11] |
Schloss P D, Westcott S L, Ryabin T, et al. Introducing mothur:Introducing mothur:Open-Source, platform-independent, communitysupported software for describing and comparing microbial communities[J]. Applied and Environmental Microbiology, 2009, 75(23): 7537-7541. DOI:10.1128/AEM.01541-09 |
| [12] |
Legendre P, Gallagher E D. Ecologically meaningful transformations for ordination of species data[J]. Oecologia, 2001, 129(2): 271-280. DOI:10.1007/s004420100716 |
| [13] |
Faith D P, Minchin P R, Belbin L. Compositional dissimilarity as a robust measure of ecological distance[J]. Vegetatio, 1987, 69(1/2/3): 57-68. |
| [14] |
Wollenberg A L V D. Redundancy analysis an alternative for canonical correlation analysis[J]. Psychometrika, 1977, 42(2): 207-219. DOI:10.1007/BF02294050 |
| [15] |
Segata N, Izard J, Waldron L, et al. Metagenomic biomarker discovery and explanation[J]. Genome Biology, 2011, 12(6): R60. |
| [16] |
Pedrós-Alió C. The rare bacterial biosphere[J]. Annual Review of Marine Science, 2012, 4: 449-466. DOI:10.1146/annurev-marine-120710-100948 |
| [17] |
陈逸斐, 韦梦, 裘琼芬, 等. 两种三疣梭子蟹(Portunus trituberculatus)养殖塘中浮游细菌的季节变化[J]. 渔业科学进展, 2015, 36(5): 119-125. CHEN Yi-fei, WEI Meng, QIU Qiong-fen, et al. The seasonal variations of bacterioplankton in two types of ponds of Portunus trituberculatus[J]. Progress in Fishery Sciences, 2015, 36(5): 119-125. |
| [18] |
范立民.吉富罗非鱼养殖池塘微生物群落研究[D].南京: 南京农业大学, 2015. FAN Li-min. Microbial community in tilapia(Oreochromis niloticus) cultural ponds[D]. Nanjing: Nanjing Agricultural University, 2015. |
| [19] |
Rosenberg E. The family Prevotellaceae[M]//Rosenberg E, DeLong EF, Lory S, et al. The prokaryotes, Berlin: Springer, 2014.
|
| [20] |
Hahn M W, Schauer M. (Candidatus Aquirestis calciphila) and (Candidatus Haliscomenobacter calcifugiens), filamentous, planktonic bacteria inhabiting natural lakes[J]. International Journal of Systematic and Evolutionary Microbiology, 2007, 57(5): 936-940. DOI:10.1099/ijs.0.64807-0 |
| [21] |
Fan L M, Barry K, Hu G D, et al. Bacterioplankton community analysis in tilapia ponds by Illumina high-throughput sequencing[J]. World Journal of Microbiology and Biotechnology, 2016, 32(1): 10. |
| [22] |
Mcgregor G B, Fabbro L D. Dominance of Cylindrospermopsis raciborskii(Nostocales, Cyanoprokaryota)in Queensland tropical and subtropical reservoirs:Implications for monitoring and management[J]. Lakes and Reservoirs:Research and Management, 2000, 5(3): 195-205. DOI:10.1046/j.1440-1770.2000.00115.x |
| [23] |
张军毅, 朱冰川.太湖(五里湖)蓝藻水华种类及其演替规律研究[C]//2012中国环境科学学会学术年会论文集(第二卷).北京: 中国环境科学出版社, 2012. ZHANG Jun-yi, ZHU Bing-chuan. A study on the species and succession of cyanobacteria bloom in Taihu Lake(Wuli Lake)[C]//Proceedings of the Annual Meeting of Chinese Society for Environmental Science, Beijing: China Environmental Press, 2012. |
| [24] |
李秋华.大镜山水库水质改善生态工程效果及浮游植物群落动态特征[D].广州: 暨南大学, 2008. LI Qiu-hua. The effects of ecotechnological engineering on improving water quality and the dynamical characteristics of phytoplankton in Dajingshan reservoir[D]. Guangzhou: Jinan University, 2008. |
| [25] |
Choi J S, Ji T K, Joo H J. Effect of total dissolved solids injection on microbial diversity and activity determined by 16S rRNA gene based pyrosequencing and oxygen uptake rate analysis[J]. Environmental Engineering Science, 2014, 31(8): 474-480. DOI:10.1089/ees.2014.0043 |
| [26] |
Adams H E, Crump B C, Kling G W. Temperature controls on aquatic bacterial production and community dynamics in arctic lakes and streams[J]. Environmental Microbiology, 2010, 12(5): 1319-1333. DOI:10.1111/j.1462-2920.2010.02176.x |
| [27] |
Tang X, Li L, Shao K, et al. Pyrosequencing analysis of free-living and attached bacterial communities in Meiliang Bay, Lake Taihu, a large eutrophic shallow lake in China[J]. Canadian Journal of Microbiology, 2015, 61(1): 22-31. |
| [28] |
Wetzel R G. Limnology:lake and river ecosystems[M]. 3rd Edition. San Diego: Academic Press, 2001.
|
| [29] |
Shade A, Jones S E, Mcmahon K D. The influence of habitat heterogeneity on freshwater bacterial community composition and dynamics[J]. Environmental Microbiology, 2008, 10: 1057-1067. DOI:10.1111/j.1462-2920.2007.01527.x |
| [30] |
Sheik C S, Beasley W H, Elshahed M S, et al. Effect of warming and drought on grassland microbial communities[J]. The ISME Journal, 2011, 5(10): 1692-1700. DOI:10.1038/ismej.2011.32 |
| [31] |
O'Neil J M, Davis T W, Burford M A, et al. The rise of harmful cyanobacteria blooms:The potential roles of eutrophication and climate change[J]. Harmful Algae, 2012, 14: 313-334. DOI:10.1016/j.hal.2011.10.027 |
| [32] |
Lindström E S, Langenheder S. Local and regional factors influencing bacterial community assembly[J]. Environmental Microbiology Reports, 2012, 4(1): 1-9. DOI:10.1111/j.1758-2229.2011.00257.x |
| [33] |
刘乾甫, 赖子尼, 王超, 等. 珠三角地区密养淡水鱼塘水体叶绿素a及水环境动态研究[J]. 中国农学通报, 2015, 31(2): 138-145. LIU Qian-fu, LAI Zi-ni, WANG Chao, et al. Studies on dynamics of chl a and water environment of intensive fishponds in Pearl River Delta[J]. Chinese Agricultural Science Bulletin, 2015, 31(2): 138-145. |
| [34] |
李志波, 宋超, 季丽, 等. 罗非鱼池塘环境因子对叶绿素a影响的通径分析[J]. 环境科学与技术, 2015, 38(12): 110-116. LI Zhi-bo, SONG Chao, JI Li, et al. Path analysis of effects of environmental factors on Chlorophyll-a in tilapia aquaculture ponds[J]. Environmental Science & Technology, 2015, 38(12): 110-116. |
| [35] |
Downing J A, Osenberg C W, Sarnelle O. Meta-analysis of marine nutrient-enrichment experiments:Variation in the magnitude of nutrient limitation[J]. Ecology, 1999, 80(4): 1157-1167. |
| [36] |
Huang W, McDowell W H, Zou X M, et al. Dissolved organic carbon in headwater streams and riparian soil organic carbon along an altitudinal gradient in the Wuyi Mountains, China[J]. PLoS One, 2013, 8(11): e78973. DOI:10.1371/journal.pone.0078973 |
| [37] |
Bai Y, Shi Q, Wen D, et al. Bacterial communities in the sediments of Dianchi Lake, a partitioned eutrophic waterbody in China[J]. PLoS One, 2012, 7(5): e37796. DOI:10.1371/journal.pone.0037796 |
| [38] |
Fierer N, Bradford M A, Jackson R B. Toward an ecological classification of soil bacteria[J]. Ecology, 2007, 88(6): 1354-1364. |