 2021, Vol. 40
2021, Vol. 40



土壤碳库的碳储量是大气碳库的3倍,大约为1 580 Gt,因此其对全球碳循环至关重要[1]。土壤有机碳参与土壤养分转化并且比较活跃,土壤生物通过活动将有机碳分解利用并释放出CO2,从而对全球碳平衡产生重大影响[2-3]。有机碳以不同形式存在于团聚体中而组成土壤结构,同时其又受团聚体保护而难被分解,二者之间的相互作用对促进土壤固碳至关重要[4]。各粒径团聚体形成的机制、孔隙特征以及土壤微生物分解其中有机碳的难易程度的差异[4-5],预示着不同粒径团聚体有机碳矿化作用释放CO2的机制有可能不同。因此,研究不同粒径团聚体中有机碳的矿化特征,分析不同粒径团聚体在全土有机碳矿化中发挥的作用,对于进一步理解有机碳在土壤中的转化、定量评价土壤碳平衡具有重要意义。
坡耕地红壤是我国重要的土壤资源,红壤区也是重要的农业生产区域,但由于人类活动频繁,使农田土壤受影响较大,加剧了水土流失的发生,制约了农业生产[6],因此也成为了许多学者的关注对象。近年来很多学者针对红壤团聚体及其有机碳进行了研究。向蕊等[1]的研究发现,绝大部分的红壤有机碳储藏在>2 mm和2~0.25 mm的团聚体中,并且在低氮水平下,间作显著增加了>2 mm团聚体的比例及其有机碳含量。李燕燕等[7]的研究发现,植被恢复模式下红壤大团聚体的比例最高,微团聚体的比例最低,大团聚体总有机碳的储量最大。谢锦升等[8]的研究发现,植被恢复能够显著增加侵蚀退化红壤大团聚体的含量及其中有机碳的比例。以上研究较多关注的仅是团聚体的分布特征和其中有机碳的分布特征,从团聚体粒径的角度考虑,也有部分学者对于红壤团聚体微域空间内有机碳的矿化作用进行了研究。陈晓芬等[9]的研究发现,不同施肥处理的红壤性水稻土的有机碳累积矿化量在>2 mm和1~2 mm团聚体中最高,在0.053~0.25 mm团聚体中最低。杨予静等[10]对南亚热带马尾松人工林土壤的研究发现,1~2 mm团聚体有机碳累积矿化量最高,原状土和<0.25 mm团聚体的易矿化碳库的矿化速率由于穿透雨减少而显著降低。但总体而言,关于红壤团聚体水平上的有机碳矿化作用的研究依然少见报道,因此需要更多相关研究来丰富对红壤团聚体有机碳矿化的认识。
合理的作物种植模式可以实现对光热水肥的高效利用,对于团聚体稳定性的提高及其中有机碳含量的提升效果显著[1, 11]。单作和间作种植模式下作物根系释放的根系分泌物的量有差异,植物残体的种类和归还数量也有差异,这些改变了土壤颗粒的结合强度,从而影响土壤团聚体的形成、稳定性和有机碳的分解速率[11]。已有的研究指出,间作可以提高土壤的有机碳含量,也可以提高团聚体有机碳含量[1]。有学者研究了不同种植模式下室内短期培养的土壤有机碳矿化动态,如贾曼莉等[12]研究了渭北苹果园清耕、苹果间作白三叶、苹果间作小冠花3种模式下土壤有机碳的矿化动态,结果表明两种间作处理的土壤有机碳矿化累积量显著高于苹果园清耕。这些研究关注的是不同种植模式下土壤有机碳和团聚体有机碳的分布特征以及在全土水平上土壤有机碳的矿化作用,而在团聚体水平上,种植模式对有机碳矿化的影响如何未见报道,还需进一步研究来揭示。
本研究以坡耕地红壤为研究对象,研究田间不同种植模式下全土及不同粒径团聚体中有机碳的矿化动态,并初步分析了不同粒径团聚体对土壤总有机碳矿化量的贡献,以进一步阐明土壤有机碳的矿化规律,为了解坡耕地红壤不同种植模式下不同粒径团聚体中有机碳转化和稳定的作用机制提供理论依据。
1 材料与方法 1.1 供试土壤和作物品种试验于2019年4—10月在云南省昆明市盘龙区松华坝水源保护区大摆社区进行(25°02' 28.8″ N,102°58'39.7″E),该地坡度为10°,海拔2 234 m,年平均气温16.0 ℃,年均降水量900~1 000 mm,降水主要集中在5—9月,属亚热带高原季风气候。供试土壤类型为红壤,质地为黏土,0~20 cm耕层土壤基本理化性质:有机质含量37.02 g·kg-1,碱解氮105.00 mg· kg-1,速效磷26.83 mg·kg-1,速效钾217.90 mg·kg-1,pH 5.20。供试玉米为"云瑞88",大豆为"滇杂7号",白萝卜为"水白晶"。
1.2 试验设计田间小区定位试验始于2018年。在同一坡面上采取随机区组设计,各小区地块规格为4 m×10 m,坡度为10°,小区与小区间均间隔1 m作为人行过道。试验共设置5个处理,分别为:玉米单作(MM)、大豆单作(MS)、白萝卜单作(MR)、玉米大豆间作(IMS)、玉米白萝卜间作(IMR),每个处理设置3个重复,共15个小区。玉米单作小区共计17行,采用宽窄行种植,宽行80 cm,窄行40 cm,每行16株,株距25 cm;单作大豆小区共计17行,采用等行距种植,行距60 cm,每行32株,株距25 cm;单作白萝卜小区共计17行,采用等行距种植,行距60 cm,每行13株,株距30 cm;间作玉米大豆采用2∶2种植模式,小区共计玉米12行,每行13株,大豆11行,玉米与玉米间、大豆与大豆间行距均为40 cm,玉米与大豆间行距50 cm,每行26株,株距均为30 cm;间作玉米白萝卜采用2∶2种植模式,小区共计玉米12行,白萝卜11行,玉米与玉米间、白萝卜与白萝卜间行距均为40 cm,玉米与白萝卜间行距50 cm,玉米每行13株,株距30 cm,白萝卜每行11株,株距35 cm。
按照当地常规施肥量,玉米施N 315 kg·hm-2、P2O5 120 kg·hm-2、K2O 120 kg·hm-2,其中氮肥50% 作基肥施用,50% 作追肥在大喇叭口期施用,磷肥和钾肥全部作基肥施用;大豆一次性施N 120 kg·hm-2、P2O5 240 kg·hm-2、K2O 180 kg·hm-2;白萝卜一次性施N 150 kg·hm-2、P2O5 150 kg·hm-2、K2O 225 kg·hm-2。所用氮肥为尿素(含N 46%),磷肥为过磷酸钙(含P2O5 16%),钾肥为硫酸钾(含K2O 51%)。玉米在播种、施肥、覆土后灌水并盖塑料薄膜,在作物生长前期和中期视土壤干旱情况和杂草生长情况适时进行灌水和除草等田间管理。
1.3 样品采集与分析于2019年10月13日采集玉米、大豆、白萝卜成熟期的土壤样品。每一个种植小区都分别随机选取有代表性(株高、茎秆粗细、长势等相近)的植株并标记好位置,待作物收获后取土样,具体步骤为:单作玉米和单作白萝卜小区分别随机选取3株有代表性的植株;单作大豆小区随机选取6株(3个点,1个点2株)有代表性的植株;玉米大豆间作小区随机选取3株有代表性的玉米和相对应的6株(3个点)有代表性的大豆;玉米白萝卜间作小区随机选取3株有代表性的玉米和相对应的3株有代表性的白萝卜。收获后以植株生长点为中心画30 cm×30 cm的正方形,在此区域内挖取2个0~20 cm原状土柱组成一个混合样品,以确保混合土壤样品具有代表性。将土壤样品储存在硬质塑料盒中并迅速运回实验室处理。用四分法选取一部分土壤在室内剔除石砾、植物残根等杂物,混合均匀后自然风干,风干土一部分供团聚体分级使用,另一部分过2 mm筛供全土有机碳的测定使用;剩余部分新鲜土壤样品过2 mm筛供全土有机碳矿化培养使用。
土壤团聚体的分级采用干筛法[13]。将预处理后的风干土壤样品混合均匀后,过5 mm筛备用。将过5 mm筛后的土壤样品置于套筛(孔径依次为2 mm、0.25 mm)顶部并安装底盒和筛盖后,以频率150次· min-1,左右振幅10 cm,振荡5 min。收集各粒径团聚体样品称质量后计算其质量分数。将收集到的各级团聚体一部分用于矿化培养试验,另一部分用于测定团聚体有机碳含量。
有机碳的测定采用重铬酸钾容量法-外加热法[14]。
矿化培养试验采用室内恒温培养,碱液吸收法测定[9]。称20.00 g新鲜全土土壤样品和3个粒径风干团聚体样品平铺于500 mL培养瓶底部,用去离子水调节全土和各粒级团聚体的含水量至田间饱和持水量的60%,密封并置于25 ℃恒温培养箱中预培养一周以恢复微生物活性,然后将盛有10 mL 0.1 mol·L-1 NaOH溶液的25 mL吸收杯悬挂于培养瓶中,密封好后在25 ℃恒温培养箱中暗培养。每个培养装置设置3个平行试验,同时设3个无土空白对照,在培养的第1、3、5、7、10、14、21、28 d进行测定,在每次测定时采用称质量的方法加水以确保土壤样品水分保持在田间持水量的60%,测定时用镊子小心更换碱液吸收杯,向换出来的吸收杯中加入1 mol·L-1 BaCl2溶液2 mL,再滴加2滴酚酞指示剂,用0.1 mol·L-1 HCl(每次滴定前用硼砂进行标定)滴定至红色消失。根据HCl滴定量计算培养期内全土和团聚体中有机碳的矿化量。
1.4 数据处理

|
式中:CHCl为盐酸浓度,mol·L-1;V0为空白处理消耗的盐酸体积,mL;V1为消耗盐酸的体积,mL。
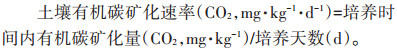
|
土壤有机碳累积矿化量指从培养开始至某一时间点土壤CO2的总释放量,采用一级动力学方程进行拟合[15],即:
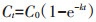
|
式中:Ct为经过t时间后土壤有机碳的累积矿化量,g· kg-1;C0为土壤潜在可矿化有机碳量,g·kg-1;k为有机碳库的周转速率常数,d-1;t为培养天数,d。半周转期T1/2=ln2/k。
各粒级团聚体有机碳矿化对全土有机碳矿化的贡献率(%)的计算公式[9]为:某一粒级团聚体有机碳矿化对全土有机碳矿化的贡献率=该粒级团聚体有机碳矿化量×该粒级团聚体含量/全土有机碳矿化量× 100%。
试验数据采用SPSS 22.0进行单因素方差分析(One-way ANOVA)和Duncan差异显著性检验(P<0.05);用Excel 2016作图,Origin Pro 2019b进行一级动力学方程拟合。图表中数据均为平均值±标准差。
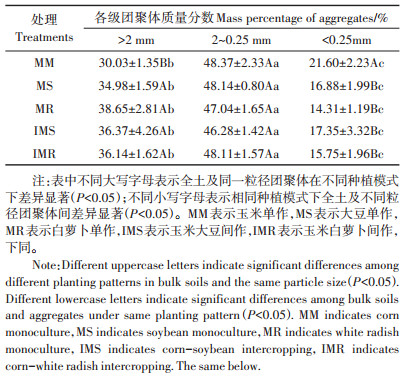
2 结果与分析 2.1 不同种植模式下土壤团聚体粒径分布从表 1可以看出,不同种植模式下团聚体粒径分布表现为2~0.25 mm团聚体含量最高,其次是>2 mm团聚体,<0.25 mm团聚体的含量最低。MM处理>2 mm团聚体的质量分数较MS、MR、IMS和IMR处理分别显著降低了4.95、8.62、6.34和6.11个百分点(P<0.05)。MM处理<0.25 mm的团聚体含量较MS、MR、IMS和IMR处理分别显著提高了0.28、0.51、0.25倍和0.37倍(P<0.05)。
|
|
表 1 不同种植模式下团聚体粒径分布 Table 1 The content distribution of soil aggregates under different planting patterns |
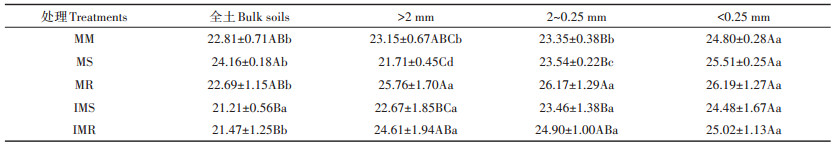
研究结果显示(表 2),在相同种植模式下,土壤有机碳在<0.25 mm团聚体中的含量高于在全土、>2 mm和2~0.25 mm团聚体的含量。在不同种植模式下,对于全土而言,MS处理的有机碳含量较IMS处理显著增加了0.14倍。在>2 mm、2~0.25 mm和<0.25 mm团聚体中,IMS和IMR处理的有机碳含量与相对应的单作处理间未达到显著差异水平(P>0.05)。
|
|
表 2 不同种植模式下全土及不同粒径土壤团聚体有机碳含量(g·kg-1) Table 2 The content of SOC in soil and soil aggregates with different diameters under different planting patterns(g·kg-1) |
全土和各粒径团聚体有机碳矿化速率均随培养时间的增加呈现出不同程度的下降趋势(图 1)。全土有机碳矿化速率在整个培养过程中呈波动式缓慢递减趋势,在培养的前14 d CO2的产生速度相对较快,5个处理峰值平均为57.49 mg·kg-1·d-1。>2 mm团聚体的有机碳矿化速率在培养后的第1 d排序为MS> IMS>MM>MR>IMR,培养结束时有机碳矿化速率平均为培养最初的30.28%。2~0.25 mm团聚体的有机碳矿化速率在培养结束时平均为培养最初的18.52%。<0.25 mm团聚体的有机碳矿化速率在开始培养后的第3 d平均为41.39 mg·kg-1·d-1,第14 d快速下降,平均为16.91 mg·kg-1·d-1,随后逐渐趋于稳定直至培养结束。在培养开始后的第1 d,2~0.25 mm团聚体有机碳矿化速率平均最高,其次为全土,再次为>2 mm团聚体,最低为<0.25 mm团聚体。在整个培养期,全土及不同粒径团聚体有机碳矿化速率随培养时间的延长不断波动变化,因此并不能明显区分不同种植模式之间的差异。

|
MM表示玉米单作,MS表示大豆单作,MR表示白萝卜单作,IMS表示玉米大豆间作,IMR表示玉米白萝卜间作。下同 MM indicates corn monoculture, MS indicates soybean monoculture, MR indicates white radish monoculture, IMS indicates corn-soybean intercropping, IMR indicates corn-white radish intercropping. The same below 图 1 不同种植模式下全土和团聚体有机碳矿化速率变化 Figure 1 Changes of mineralization rate of organic carbon in bulk soils and aggregate fractions under different planting patterns |
不同种植模式下,培养28 d时全土和团聚体有机碳累积矿化量情况见图 2。在相同的种植模式下,全土及不同粒径团聚体有机碳累积矿化量整体表现为全土最高,其次为2~0.25 mm团聚体,再次为<0.25 mm团聚体,>2 mm团聚体最低。不同种植模式下,对于全土,MS处理的有机碳累积矿化量较MM和IMS处理分别显著增加了14.09% 和6.97%,IMS处理较MM处理显著增加了6.66%(P<0.05)。对于>2 mm团聚体,MS处理的有机碳累积矿化量较IMS处理显著增加了25.21%,MM处理较IMR处理显著增加了6.00% (P<0.05)。对于2~0.25 mm团聚体,MS处理的有机碳累积矿化量较IMS处理显著增加了9.84%,MM处理的有机碳累积矿化量较IMS和IMR处理分别显著增加了6.34%和21.36%(P<0.05)。对于<0.25 mm团聚体,各处理间有机碳累积矿化量差异均不显著。

|
不同大写字母表示全土及同一粒径团聚体在不同种植模式下差异显著(P<0.05);不同小写字母表示相同种植模式下全土及不同粒径团聚体间差异显著(P<0.05) Different uppercase letters indicate significant differences among different planting patterns in bulk soils and the same particle size(P<0.05).Different lowercase letters indicate significant differences among bulk soils and aggregates under same planting pattern(P<0.05) 图 2 培养28 d时不同种植模式下全土及团聚体有机碳累积矿化量 Figure 2 Cumulative amounts of organic carbon mineralization in bulk soils and aggregate fractions under different planting patterns in 28 days |
本研究土壤及团聚体有机碳累积矿化量与培养天数之间的动态变化采用一级动力学方程Ct =C0(1- e-kt) 进行拟合(P<0.01),且该模型的拟合效果较好,决定系数R2>0.99(表 3)。结果表明,在相同种植模式下,C0和C0/SOC值在全土中最高,且显著高于其他3个粒径团聚体(P<0.05)。<0.25 mm团聚体的C0和C0/ SOC值最低,2~0.25 mm团聚体的k值最高,>2 mm团聚体的T1/2最高。在不同种植模式下,MS处理的C0在全土和3个粒径团聚体中都显著高于IMS处理(P<0.05)。在全土、>2 mm团聚体和<0.25 mm团聚体中,IMS处理的k值都显著高于MM处理(P<0.05)。对于>2 mm和<0.25 mm团聚体,MM和MS处理的T1/2显著高于IMS处理,在2~0.25 mm团聚体中,IMS和IMR处理的T1/2分别较MM处理显著提高了6.75%和10.74% (P<0.05)。另外,在全土中,MM处理的C0/SOC值较IMR处理显著提高了18.53%,IMR处理较MR处理显著提高了13.41%(P<0.05)。对于>2mm团聚体,MM处理的C0/SOC值较IMS和IMR处理分别显著提高了50.93%和13.02%(P<0.05)。在2~0.25mm团聚体中,MM处理的C0/SOC值较IMR处理显著提高了14.25% (P<0.05)。
|
|
表 3 培养28 d时土壤及团聚体矿化动力学方程参数 Table 3 The parameters of kinetic equations of bulk soils and aggregate fractions after 28 days of incubation |

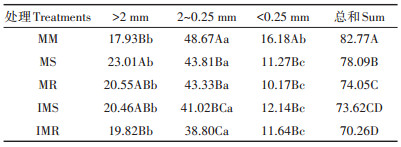
研究结果显示(表 4),各粒径团聚体有机碳矿化对全土有机碳矿化贡献率的总和在各处理中依次为MM (82.77%)>MS(78.09%)>MR(74.05%)>IMS(73.62%)> IMR(70.26%)。2~0.25 mm团聚体对全土有机碳矿化的贡献率最高,范围在38.80%~48.67%,其次是>2 mm团聚体,范围在17.93%~23.01%,贡献率最低的是<0.25 mm团聚体,范围在10.17%~16.18%。
|
|
表 4 不同种植模式下各粒径团聚体对全土有机碳矿化的贡献率(%) Table 4 Contributions of aggregate fractions to soil organic carbon mineralization under different planting patterns(%) |
土壤团聚体是土壤结构的基本单元,其数量和质量决定土壤理化性状及其协调状况[4]。本研究对团聚体的分级采用干筛法,因为干筛法较湿筛法降低了水分张力等因素对土壤结构的干扰,减少了对土壤中某些易变性有机胶结物质的破坏,也减少了对团聚体中微生物群落物理生境的破坏,对于旱地红壤,干筛分离出的团聚体更接近田间实际[16]。本研究中,各种植模式下>2 mm和2~0.25 mm团聚体含量较高,这与张鹏等[17]的研究结果相似。这是因为铁铝氧化物是南方红壤地区微团聚体的主要胶结物质,70%以上的土壤有机碳存在于粉-黏团聚体中,而大团聚体的主要胶结物质是有机质,有机质将微团聚体胶结成大团聚体[1, 4],并且由于新鲜有机物质的输入,作物根系、土壤动物、微生物及他们的代谢产物增多,同时土壤中有机胶结物质和载体介质的量也增多[18],从而促进了>0.25 mm团聚体的形成。本研究进一步发现,MM处理中>2 mm团聚体含量最低,而<0.25 mm团聚体含量最高(P<0.05),这可能是因为玉米丰富的根系对土壤的扰动更大,大颗粒团聚体中的颗粒有机物有助于微团聚体的形成,由于玉米根系的扰动和颗粒有机物的分解,大颗粒团聚体破碎使得微团聚体被释放出来[1, 19],因此大团聚体比例减少,微团聚体比例增加。
有机碳的含量是有机物质输入和矿化分解动态平衡的结果[20]。本研究发现,在相同种植模式下<0.25 mm团聚体的有机碳含量最高(P<0.05)。这是因为微团聚体内的孔隙极小,微生物分解有机碳耗能很多,从而使有机碳的分解降低,微团聚体中的有机碳受到的干扰较少,所以储存的时间更长[4]。前人的研究发现,玉米马铃薯[1]、苹果白三叶[18]、蚕豆玉米、小麦玉米、小麦蚕豆[21]的间作体系均能提高土壤有机碳含量。本研究结果与上述研究结果不同,本研究结果表明,IMS使全土的有机碳含量呈现亏损的状态(P<0.05),其他各粒径团聚体有机碳含量在单作与间作之间并没有显著差异,间作可提高有机质的相对分解速率,但有机质快速分解并未使土壤有机碳含量得到提高[22]。间作系统的根系发达,凋落物更具多样性,土壤微生物也增多,这使得碳分解大于碳输入[23-24]。另外,本研究结果与种植年限短有关,上述研究结果是基于多年的田间定位试验,而本研究所在的田间小区定位试验始于2018年,土壤还具有较好的缓冲能力,这使得有机碳含量因种植模式发生变化具有了一定的滞后性,所以本研究中单作与间作模式团聚体有机碳含量没有显著差异,这还需要通过进一步的长期定位试验来验证。
土壤有机碳矿化是土壤碳库平衡的重要过程,其速率的大小表示有机碳分解的快慢程度[9],对其进行研究有助于揭示土壤碳库的动态机理[3]。本研究中,团聚体有机碳矿化与全土有机碳矿化的变化规律一致。总体上,全土及不同粒径团聚体在培养前期的有机碳矿化速率大,并且呈现快速下降趋势,后期有机碳矿化速率,随培养时间推移逐渐减慢并保持相对稳定,这与土壤微生物活性和有机碳矿化难易程度有关。有机碳是由容易矿化和不易矿化两部分组成的,室内培养28 d所分解的有机碳多为易矿化有机碳。培养前期,土壤及团聚体样品加水到田间持水量的60%激发了微生物的活性,土壤中容易矿化的活性有机碳(如糖类和蛋白质等)较多,微生物将其迅速分解利用后释放出大量的CO2,因此有机碳矿化速率较高;培养时间越长,土壤中的活性有机碳被消耗得越多,微生物可利用的有效碳源越来越少,所以有机碳矿化速率逐渐降低并表现出相对稳定的趋势[15, 25]。
土壤有机碳矿化与团聚体密切相关,团聚体粒径的大小能够影响土壤有机碳的矿化速率,进而影响土壤有机碳的累积矿化量[5, 9]。本研究中,除全土外,2~ 0.25 mm团聚体有机碳累积矿化量最高(P<0.05),这是由于2~0.25 mm团聚体稳定性低,其含有的活性有机碳较多并且易分解[4-5],有机碳矿化速率较高,因此其累积矿化量也高。本研究进一步发现,全土中IMS处理的有机碳累积矿化量显著高于MM(P<0.05),这可能与凋落物的性质和数量有关。有研究[26]表明凋落物进入土壤后可以促进有机碳的矿化,由于凋落物作为外源有机物质输入到土壤中的有机碳的质量和数量不同,从而使含有不同凋落物的土壤有机碳矿化也不同[27-28]。贾曼莉等[12]对渭北生草果园土壤有机碳矿化的研究表明,果园生草间作显著提升了矿化率,证实了上述观点。除此之外,本研究还发现,在全土、>2 mm团聚体和2~0.25 mm团聚体中,与间作处理相比,有机碳累积矿化量在某些单作处理中显著提高(P<0.05),并且在MS处理中有机碳累积矿化量最高。产生这种不同差异的原因很复杂,除了受作物凋落物的性质和数量的影响外[27-28],土壤微生物活性、种类和数量的不同[29-30],作物根系以及根系分泌物的不同也都会影响有机碳的含量和稳定性[11]。本课题组前期[11]的研究发现,间作会促进根系总糖和总有机酸的分泌,增强根际细菌的活力,促进多糖分泌与菌丝生长,对微团聚体的胶黏作用和缠绕作用也得到增强,这些有利于微团聚体转化为大颗粒的团聚体,使间作更能储存碳,因此,使得单作处理比间作处理的有机碳矿化作用更强。
本研究利用一级动力学方程拟合了土壤和团聚体有机碳矿化动态过程,其中,C0表示在理想情况下有机碳矿化释放的CO2的量,C0/SOC值则表示土壤有机碳矿化分解作用消耗土壤中有机碳的比例,可反映土壤固碳能力的强弱,该比例越低,表明土壤的固碳能力越强,反之,则固碳能力越弱[31]。本研究中,在相同种植模式下,<0.25 mm团聚体的C0和C0/SOC值最低(P<0.05),说明<0.25 mm团聚体较其他两种粒径团聚体的固碳能力更强。这是由于团聚体团聚过程中胶结物质的差异,使有机碳受到的物理保护程度各异。大粒径团聚体中是易分解的瞬变胶结有机物,微团聚体中则多是持久性胶结剂[32]。大团聚体中含有较多易分解的活性有机碳,微团聚体中则含有高度腐殖化的惰性有机碳[33]。本研究还发现,在相同种植模式下,>2 mm团聚体的T1/2最高(P<0.05)。这是由于大团聚体与空气接触的表面积小,其表面的有机碳(包括有机胶结剂)被分解的几率也小,而且大团聚体形成后其内部的孔隙降低,孔隙间的空气和水分相应减少,使有机碳与矿物颗粒的结合更加紧密,从而降低了其中有机碳的分解[4, 32]。另外,本研究中2~0.25 mm团聚体有机碳库的k值在相同种植模式下最高(P<0.05)。这是由于2~0.25 mm大团聚体颗粒组中聚集了大量易氧化态有机碳,所以该粒径团聚体中有机碳稳定性较低,周转快[34-35]。
全土有机碳矿化表现出不同粒径团聚体有机碳矿化的总体特征。本研究中,2~0.25 mm团聚体对全土有机碳矿化的贡献最大,其次是>2 mm团聚体,<0.25 mm团聚体对全土有机碳矿化的贡献最小,这与陈晓芬等[9]在红壤性水稻土中发现的>0.25 mm大团聚体对全土有机碳矿化的贡献高的研究结果相类似。有学者研究证明大粒径团聚体对土壤碳排放的贡献最大与大粒径团聚体含量占主要优势有关[5]。综上所述,大粒径团聚体在土壤有机碳矿化中是发挥主导作用的。另外,本研究中对全土的矿化培养采用的是鲜土,团聚体的矿化培养采用的是风干土,即使有1周的预培养缓冲期以恢复微生物活性,风干土壤微生物量、微生物群落结构以及代谢能力等也会发生变化[36],这可能是导致各粒径团聚体对全土有机碳矿化的贡献率总和不高的原因。
由于试验条件的限制,本研究采用碱液吸收法在室内恒温条件下模拟不同种植模式下坡耕地红壤团聚体有机碳的矿化特征,采用一级动力学方程拟合矿化过程并根据拟合出的相关参数在一定程度上预测田间土壤有机碳矿化的动态变化趋势。但是在田间条件下影响土壤有机碳矿化的环境因素非常复杂,如水分、温度、土壤受扰动程度等都会影响土壤有机碳矿化,实验室培养过程是在培养瓶中进行,通气状况、温度、水分等与田间实际环境有所不同,从而会对有机碳的矿化过程产生影响。因此,本研究结果与田间条件下多因素综合作用的结果可能存在差异,今后仍需加强相关控制试验的研究以及土壤有机碳矿化的田间原位试验研究。
4 结论(1) 不同种植模式下2~0.25 mm团聚体含量最高,<0.25 mm团聚体含量最低。玉米单作后土壤>2 mm团聚体比例有所下降,<0.25 mm团聚体比例有所增加。在相同种植模式下,<0.25 mm团聚体有机碳含量显著高于全土、>2 mm团聚体和2~0.25 mm团聚体。全土中玉米大豆间作处理的土壤有机碳含量显著低于大豆单作处理,不同粒径团聚体中有机碳含量在单作处理与相对应间作处理之间没有显著差异。
(2) 不同种植模式下全土与各粒径团聚体的有机碳矿化速率均随培养时间的增加呈现不同程度的下降趋势,培养前期波浪式递减,培养后期逐渐趋于稳定。总体上,不同种植模式下全土和团聚体中有机碳矿化作用的强弱大致表现为单作处理比相对应的间作处理更强,并且大豆单作处理的有机碳矿化作用最强。在相同种植模式下,2~0.25 mm团聚体中的有机碳矿化速率最快、更易矿化,因此其有机碳累积矿化量显著高于其他两种粒径团聚体。<0.25 mm团聚体的潜在可矿化有机碳量与C0/SOC值最低,说明其固碳能力最强。>2 mm和2~0.25 mm团聚体对全土有机碳矿化的贡献最大。
| [1] |
向蕊, 伊文博, 赵薇, 等. 间作对土壤团聚体有机碳储量的影响及其氮调控效应[J]. 水土保持学报, 2019, 33(5): 303-308. XIANG R, YI W B, ZHAO W, et al. Effects of intercropping on soil aggregate-associated organic carbon storage and nitrogen regulation[J]. Journal of Soil and Water Conservation, 2019, 33(5): 303-308. |
| [2] |
吕真真, 刘秀梅, 仲金凤, 等. 长期施肥对红壤性水稻土有机碳矿化的影响[J]. 中国农业科学, 2019, 52(15): 2636-2645. LÜ Z Z, LIU X M, ZHONG J F, et al. Effects of long-term fertilization on mineralization of soil organic carbon in red paddy soil[J]. Scientia Agricultura Sinica, 2019, 52(15): 2636-2645. DOI:10.3864/j.issn.0578-1752.2019.15.008 |
| [3] |
CAO M, WOODWARD F I. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change[J]. Nature, 1998, 393(6682): 249-252. DOI:10.1038/30460 |
| [4] |
刘中良, 宇万太. 土壤团聚体中有机碳研究进展[J]. 中国生态农业学报, 2011, 19(2): 447-455. LIU Z L, YU W T. Review of researches on soil aggregate and soil organic carbon[J]. Chinese Journal of EcoAgriculture, 2011, 19(2): 447-455. |
| [5] |
王菁, 陈防, 刘毅. 不同粒级土壤团聚体呼吸特征及其对碳排放的贡献[J]. 植物科学学报, 2014, 32(6): 586-593. WANG J, CHEN F, LIU Y. Respiration characteristics of different sized soil aggregates and their contribution to carbon emissions[J]. Plant Science Journal, 2014, 32(6): 586-593. |
| [6] |
成艳红, 武琳, 孙慧娟, 等. 稻草覆盖和香根草篱对红壤水稳性团聚体组成及有机碳含量的影响[J]. 生态学报, 2016, 36(12): 3518-3524. CHENG Y H, WU L, SUN H J, et al. Effects of straw mulching and Vetiver grass hedgerows on the size distribution of the soil water stable aggregates and aggregate-associated organic carbon in red soil[J]. Acta Ecologica Sinica, 2016, 36(12): 3518-3524. |
| [7] |
李燕燕, 刘亮英, 张志坚, 等. 亚热带红壤区自然恢复草地转换为人工林后对土壤团聚体有机碳周转的影响[J]. 水土保持学报, 2019, 33(1): 80-85. LI Y Y, LIU L Y, ZHANG Z J, et al. Effects of conversion of degraded grassland into plantations on organic carbon turnover of soil aggregate in subtropical red soil area[J]. Journal of Soil and Water Conservation, 2019, 33(1): 80-85. |
| [8] |
谢锦升, 杨玉盛, 陈光水, 等. 植被恢复对退化红壤团聚体稳定性及碳分布的影响[J]. 生态学报, 2008, 28(2): 702-709. XIE J S, YANG Y S, CHEN G S, et al. Effects of vegetation restoration on water stability and organic carbon distribution in aggregates of degraded red soil in subtropics of China[J]. Acta Ecologica Sinica, 2008, 28(2): 702-709. DOI:10.3321/j.issn:1000-0933.2008.02.031 |
| [9] |
陈晓芬, 刘明, 江春玉, 等. 不同施肥处理红壤性水稻土团聚体有机碳矿化特征[J]. 中国农业科学, 2018, 51(17): 3325-3334. CHEN X F, LIU M, JIANG C Y, et al. Organic carbon mineralization in aggregate fractions of red paddy soil under different fertilization treatments[J]. Scientia Agricultura Sinica, 2018, 51(17): 3325-3334. DOI:10.3864/j.issn.0578-1752.2018.17.008 |
| [10] |
杨予静, 刘世荣, 陈琳, 等. 马尾松人工林土壤和团聚体有机碳矿化对穿透雨减少的响应[J]. 应用生态学报, 2018, 29(6): 1779-1786. YANG Yu-jing, LIU S R, CHEN L, et al. Responses of soil and aggregate-associated organic carbon mineralization to throughfall reduction in a Pinus massoniana plantation[J]. Chinese Journal of Applied Ecology, 2018, 29(6): 1779-1786. |
| [11] |
王婷, 李永梅, 王自林, 等. 间作对玉米根系分泌物及团聚体稳定性的影响[J]. 水土保持学报, 2018, 32(3): 185-190. WANG T, LI Y M, WANG Z L, et al. Effects of intercropping on maize root exudates and soil aggregate stability[J]. Journal of Soil and Water Conservation, 2018, 32(3): 185-190. |
| [12] |
贾曼莉, 郭宏, 李会科. 渭北生草果园土壤有机碳矿化及其与土壤酶活性的关系[J]. 环境科学, 2014, 35(7): 2777-2784. JIA M L, GUO H, LI H K. Mineralization of soil organic carbon and its relationship with soil enzyme activities in apple orchard in Weibei[J]. Environmental Science, 2014, 35(7): 2777-2784. |
| [13] |
李景. 高碳氮投入农田生态系统土壤团聚体有机碳及微生物特性研究[D]. 北京: 中国农业大学, 2018. LI J. Response of soil organic carbon and microbial properties within aggregates to high-input fertilizer practices under an intensive agricultural system[D]. Beijing: China Agricultural University, 2018. |
| [14] |
鲍士旦. 土壤农业化学分析[M]. 3版. 北京: 中国农业出版社, 1999: 30-34. BAO S D. Analytical methods for soil and agricultual chemistry[M]. Third edition. Beijing: China Agriculture Press, 1999: 30-34. |
| [15] |
王朔林, 杨艳菊, 王改兰, 等. 长期施肥对栗褐土有机碳矿化的影响[J]. 植物营养与肥料学报, 2016, 22(5): 1278-1285. WANG S L, YANG Y J, WANG G L, et al. Effect of long-term fertilization on organic carbon mineralization of cinnamon soil[J]. Plant Nutrition and Fertilizer Science, 2016, 22(5): 1278-1285. |
| [16] |
CHEN L J, ZHANG S X, LIANG W J, et al. Effects of conservation tillage on soil aggregation and aggregate binding agents in black soil of Northeast China[J]. Soil & Tillage Research, 2012, 124: 196-202. |
| [17] |
张鹏, 周泉, 黄国勤. 冬季不同种植模式对稻田土壤团聚体及其有机碳的影响[J]. 核农学报, 2019, 33(12): 2430-2438. ZHANG P, ZHOU Q, HUANG G Q. Effects of winter different planting patterns on soil aggregates and organic carbon in paddy fields[J]. Journal of Nuclear Agricultural Sciences, 2019, 33(12): 2430-2438. DOI:10.11869/j.issn.100-8551.2019.12.2430 |
| [18] |
王英俊, 李同川, 张道勇, 等. 间作白三叶对苹果/白三叶复合系统土壤团聚体及团聚体碳含量的影响[J]. 草地学报, 2013, 21(3): 485-493. WANG Y J, LI T C, ZHANG D Y, et al. Effects of intercropping white clover on soil aggregates and soil organic carbon of aggregates in apple-white clover intercropping system[J]. Acta Agrestia Sinica, 2013, 21(3): 485-493. |
| [19] |
SIX J, ELLIOTT E T, PAUSTIAN K. Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture[J]. Soil Biology and Biochemistry, 2000, 32(14): 2099-2103. DOI:10.1016/S0038-0717(00)00179-6 |
| [20] |
胡雪寒, 刘娟, 姜培坤, 等. 亚热带森林转换对不同粒径土壤有机碳的影响[J]. 土壤学报, 2018, 55(6): 1485-1493. HU X H, LIU J, JIANG P K, et al. Effects of change in forest type on soil organic carbon in soil particles relative to size in subtropical China[J]. Acta Pedologica Sinica, 2018, 55(6): 1485-1493. |
| [21] |
CONG W F, HOFFLAND E, LI L, et al. Intercropping enhances soil carbon and nitrogen[J]. Global Change Biology, 2015, 21(4): 1715-1726. DOI:10.1111/gcb.12738 |
| [22] |
BOOTH M S, STARK J M, RASTETTER E. Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data[J]. Ecological Monographs, 2005, 75(2): 139-157. DOI:10.1890/04-0988 |
| [23] |
LI L, SUN J, ZHANG F. Intercropping with wheat leads to greater root weight density and larger below-ground space of irrigated maize at late growth stages[J]. Soil Science and Plant Nutrition, 2011, 57(1): 61-67. DOI:10.1080/00380768.2010.548307 |
| [24] |
CARDINALE B J, MATULICH K L, HOOPER D U, et al. The functional role of producer diversity in ecosystems[J]. American Journal of Botany, 2011, 98(3): 572-592. DOI:10.3732/ajb.1000364 |
| [25] |
吴萌, 李忠佩, 冯有智, 等. 长期施肥处理下不同类型水稻土有机碳矿化的动态差异[J]. 中国农业科学, 2016, 49(9): 1705-1714. WU M, LI Z P, FENG Y Z, et al. Dynamic differences of organic carbon mineralization in different types of paddy soil under long-term located fertilization[J]. Scientia Agricultura Sinica, 2016, 49(9): 1705-1714. |
| [26] |
史学军, 潘剑君, 陈锦盈, 等. 不同类型凋落物对土壤有机碳矿化的影响[J]. 环境科学, 2009, 30(6): 1832-1837. SHI X J, PAN J J, CHEN J Y, et al. Effects of different types of litters on soil organic carbon mineralization[J]. Environmental Science, 2009, 30(6): 1832-1837. DOI:10.3321/j.issn:0250-3301.2009.06.046 |
| [27] |
CHOW A T, TANJI K K, GAO S, et al. Temperature, water content and wet-dry cycle effects on DOC production and carbon mineralization in agricultural peat soils[J]. Soil Biology and Biochemistry, 2006, 38(3): 477-488. DOI:10.1016/j.soilbio.2005.06.005 |
| [28] |
KADONO A, FUNAKAWA S, KOSAKI T. Factors controlling mineralization of soil organic matter in the Eurasian steppe[J]. Soil Biology and Biochemistry, 2008, 40(4): 947-955. DOI:10.1016/j.soilbio.2007.11.015 |
| [29] |
汪洋, 杨殿林, 王丽丽, 等. 农田管理措施对土壤有机碳周转及微生物的影响[J]. 农业资源与环境学报, 2020, 37(3): 340-352. WANG Y, YANG D L, WANG L L, et al. Effects of farmland management measures on soil organic carbon turnover and microorganisms[J]. Journal of Agricultural Resources and Environment, 2020, 37(3): 340-352. |
| [30] |
BALSER T C, FIRESTONE M K. Linking microbial community composition and soil processes in a California annual grassland and mixed-conifer forest[J]. Biogeochemistry, 2005, 73(2): 395-415. DOI:10.1007/s10533-004-0372-y |
| [31] |
汤洁, 刘禹晴, 王思宁, 等. 吉林西部盐碱地区稻田土壤有机碳矿化特征[J]. 水土保持学报, 2019, 33(2): 162-168. TANG J, LIU Y Q, WANG S N, et al. Characteristics of organic carbon mineralization in saline-alkali paddy field in western Jilin Province[J]. Journal of Soil and Water Conservation, 2019, 33(2): 162-168. |
| [32] |
徐香茹, 汪景宽. 土壤团聚体与有机碳稳定机制的研究进展[J]. 土壤通报, 2017, 48(6): 1523-1529. XU X R, WANG J K. A review on different stabilized mechanisms of soil aggregates and organic carbon[J]. Chinese Journal of Soil Science, 2017, 48(6): 1523-1529. |
| [33] |
李景, 吴会军, 武雪萍, 等. 长期保护性耕作提高土壤大团聚体含量及团聚体有机碳的作用[J]. 植物营养与肥料学报, 2015, 21(2): 378-386. LI J, WU H J, WU X P, et al. Impact of long-term conservation tillage on soil aggregate formation and aggregate organic carbon contents[J]. Plant Nutrition and Fertilizer Science, 2015, 21(2): 378-386. |
| [34] |
周萍, 宋国菡, 潘根兴, 等. 南方三种典型水稻土长期试验下有机碳积累机制研究Ⅰ. 团聚体物理保护作用[J]. 土壤学报, 2008, 45(6): 1063-1071. ZHOU P, SONG G H, PAN G X, et al. SOC accumulation in three major types of paddy soils under long-term agro-ecosystem experiments from south China Ⅰ. Physical protection in soil micro-aggregates[J]. Acta Pedologica Sinica, 2008, 45(6): 1063-1071. DOI:10.3321/j.issn:0564-3929.2008.06.008 |
| [35] |
杨长明, 欧阳竹, 杨林章, 等. 农业土地利用方式对华北平原土壤有机碳组分和团聚体稳定性的影响[J]. 生态学报, 2006, 26(12): 4148-4155. YANG C M, OUYANG Z, YANG L Z, et al. Organic carbon fractions and aggregate stability in an aquatic soil as influenced by agricultural land uses in the Northern China Plain[J]. Acta Ecologica Sinica, 2006, 26(12): 4148-4155. DOI:10.3321/j.issn:1000-0933.2006.12.030 |
| [36] |
陈雅涵, 谢宗强. 保存过程对土壤生化指标的影响及保存土样的应用[J]. 土壤学报, 2018, 55(4): 783-796. CHEN Y H, XIE Z Q. Effects of storage on soil biochemical properties and application of stored soil[J]. Acta Pedologica Sinica, 2018, 55(4): 783-796. |