 2022, Vol. 41
2022, Vol. 41



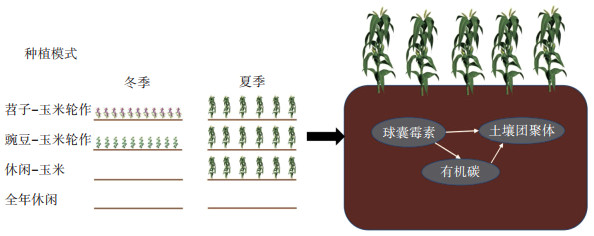
土壤侵蚀已经成为农业用地和农业生态系统一个很严重的全球性问题,土壤侵蚀对土壤质量有负面影响,进而降低作物生产力[1]。在我国云南省,由于降雨和特殊的地质地貌,坡耕地常年受到水力侵蚀的危害,特别是近些年农民对耕地的使用强度和频率越来越高,导致耕地产生土壤侵蚀的风险大幅增加,同时对土壤质量、粮食安全以及农业可持续发展产生了很大的威胁,因此,针对云南高原地区坡耕地开展土壤抗蚀机制的研究将有助于改善这一状况。
土壤团聚体是影响土壤侵蚀的重要物理因素,土壤团聚体受物理、化学和生物因素的复杂动态影响。土壤有机碳(SOC)不仅参与土壤中很多生化过程,还在团聚体形成过程中充当胶结剂[2],团聚体不仅可以为SOC提供物理保护的作用,较高的团聚体稳定性还能增强土壤中水的流动并提高储存水的能力[3],降低土壤的可蚀性。球囊霉素相关土壤蛋白(GRSP)可以作为评价土壤侵蚀程度的指标之一[4],其是由丛枝菌根真菌通过菌丝释放到土壤中的一种疏水性糖蛋白,根据提取方法的不同可分为易提取球囊霉素(EEGRSP)和总球囊霉素(T-GRSP)。大量研究发现,GRSP存在于林地[5]、耕地[6]、牧场[7]、果园[8]、休耕地[9-10]等不同土地利用方式中,而且GRSP是土壤结构维持和形成的主要成分[11],可以改善土壤团聚体稳定性,并对SOC有重要贡献,占SOC的5%~13%[9, 12]。因此可以看出,GRSP、SOC和团聚体之间关系密切。
在农业生态系统中,土壤中SOC会随着耕作和种植作物大量流失,但不同的耕作频率和种植模式会影响SOC流失的多少和速率[13]。GRSP的含量也会受到施肥[14]、耕作方式[15]、种植模式[16]、作物残留管理[6]等不同农艺措施的影响。目前国内外关于不同轮作休耕模式对GRSP分布特征的研究鲜见报道,因此,本研究以滇中坡耕地红壤作为研究对象,开展不同轮作模式和休耕等处理的田间试验,探讨GRSP、SOC和团聚体稳定性对不同种植模式的响应,并应用结构方程模型构建并分析三者之间的路径关系,以期为农业生产中土壤碳的动态过程及耕地土壤侵蚀提供一定的理论基础。
1 材料与方法 1.1 试验地概况试验地位于云南省昆明市盘龙区松华坝水源区大摆社区(25°02′ 28.8″ N,102°58′ 39.7″ E),坡度为10°,属亚热带季风气候,海拔2 234 m,年降水量900~ 1 000 mm,年平均气温16 ℃。该试验地土壤基本理化性质为:pH 6.29,有机质30.15 g·kg-1,全氮1.14 g· kg-1,碱解氮115.3 mg·kg-1,速效磷6.41 mg·kg-1,速效钾92.36 mg·kg-1。
1.2 试验设计供试作物:玉米品种为云瑞“88”;苕子和豌豆分别为当地普遍种植的毛叶紫花苕和白花矮茎豌豆。当地典型轮作模式主要为苕子/豌豆与玉米/烤烟/马铃薯轮作模式,本试验采用的夏季作物为玉米。
试验小区面积为4 m×5 m,坡度为10°。如表 1所示,试验设4个处理,每个处理3次重复,共12个小区,区组内小区随机排列。玉米采用宽窄行种植,宽行80 cm,窄行40 cm,株距25 cm。播种模式为穴播,种植时每穴两粒,待出苗后保留其中一株,单个小区植株宽行共计4行,窄行共计9行,每行16株,每个小区保持一致。苕子和豌豆采用撒播种植方式,将种子均匀撒于种完玉米的小区中,然后结合中耕和除草微翻动表土,播种深度(覆土厚度)为2~3 cm。玉米施肥采取单株精准施肥模式,施N 315 kg·hm-2,P2O5 120 kg·hm-2,K2O 120 kg·hm-2,其中氮肥分两次施用(50%基肥,50%追肥),追肥在喇叭口期进行,磷肥和钾肥全部作基肥施用;苕子和豌豆作为绿肥作物,不作施肥处理。玉米在播种、施肥、灌水后采用塑料透明薄膜覆盖。在作物各生长期内,根据作物长势情况,适时进行灌水。为避免扰动土壤,原则上不做除草工作。在冬季绿肥作物苕子和豌豆成熟后对其进行翻压还田处理;夏季玉米成熟后一部分用于测产,另一部分则将玉米收获后的秸秆还田。
|
|
表 1 试验设计及处理 Table 1 Experimental design and treatment |
在玉米成熟期采用“S”型取样方法进行土壤样品采集,每个小区采集耕作层土样,在采样和运输过程中避免挤压和扰动,保持土壤的原状结构,以免破坏团聚体。将土样按其自然结构掰成直径约5 cm的土块,剔除石块和植物根系等杂物,实验室条件下风干,用于土壤各项指标的测定。
1.3.2 指标测定团聚体测定及指标计算:采用Elliott团聚体湿筛法[17]测定>2、0.5~2、0.25~0.5 mm和 < 0.25 mm 4个粒级的水稳性团聚体。
平均质量直径(MWD):
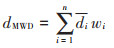
|
(1) |
式中:dMWD为平均质量直径,mm;di为各粒级水稳性团聚体平均直径,mm;wi为各粒级水稳性团聚体质量百分数,%。
GRSP的提取与测定:按照WRIGHT等[18]提出的方法提取并测定土壤中EE-GRSP和T-GRSP的含量。
总有机碳及可溶性有机碳测定[19]。总有机碳(SOC)的测定:首先进行土壤酸化,再采用multi N/C 3100 TOC分析仪进行测定。可溶性有机碳(DOC)的测定[19]:称取过2 mm筛的新鲜土样20 g于100 mL离心管中,加入40 mL蒸馏水(水土比为2∶1),在25 ℃条件下,以250 r·min-1的速度振荡0.5 h,然后于8 000 r·min-1离心10 min,上部悬浮液过0.45 μm滤膜,滤液测定前需在-18 ℃冷冻保存,滤液中有机碳含量采用multi N/C 3100 TOC分析仪测定。
1.4 数据分析采用Microsoft Excel和R语言进行数据处理和相关图表绘制。数据测定结果均以平均值±标准差的形式表示,利用单因素方差分析(One-way ANOVA)评价不同处理间各个指标的差异显著性。采用Amos软件作结构方程模型分析,以模拟GRSP对SOC含量及团聚体稳定性的影响,以及SOC对团聚体稳定性的影响。
2 结果与分析 2.1 不同种植模式下土壤团聚体组成及稳定性特征不同种植模式下水稳性团聚体组成及稳定性指标如表 2所示,4种种植模式均以0.5~2 mm粒级含量最高,其次为>2 mm粒级,这两个粒级占总团聚体的56.50%~74.06%。F-F和F-C的>2 mm粒级的团聚体含量显著高于V-C(P < 0.05),F-F的0.5~2 mm粒级团聚体含量显著高于F-C(P < 0.05),0.25~0.5 mm粒级团聚体各处理间无显著差异,对于 < 0.25 mm的微团聚体和黏粉粒,V-C中的含量显著高于其他处理(P < 0.05),F-C和P-C显著高于F-F(P < 0.05)。从≥0.25 mm粒级团聚体来看,F-F、F-C和P-C分别比V-C显著提高20.03%、10.52%和9.68%(P < 0.05),对于MWD值,F-F和F-C分别比V-C显著高30.43% 和17.39%(P < 0.05),P-C比V-C高15.22%,但两者差异不显著。因此,F-F处理的团聚体稳定性最高,V-C最低,F-C和P-C之间差异不显著,说明与V-C相比,全年休闲能较大程度地提高团聚体稳定性,而F-C也能提高团聚体稳定性,但效果不及F-F。
|
|
表 2 不同种植模式下团聚体组成及稳定性指标 Table 2 Aggregate composition and stability indicators under different planting patterns |

不同种植模式下土壤各团聚体粒级中EEGRSP、T-GRSP、SOC和DOC的含量如表 3所示。不同处理土壤中EE-GRSP的含量为0.15~0.32 mg·g-1,其中,与F-C相比,F-F处理在>2、0.5~2、0.25~0.5 mm和 < 0.25 mm 4个粒级下分别显著提高71.35%、78.61%、79.87% 和90.54%(P < 0.05);V-C处理在4个粒级下分别显著提高45.95%、58.96%、53.89% 和70.95%(P < 0.05);P-C处理在4个粒级下分别提高34.59%、45.09%、38.96% 和39.19%,且在0.5~2 mm粒级下差异显著(P < 0.05)。不同处理土壤中T-GRSP的含量为1.22~1.88 mg·g-1,其中,与F-C相比,F-F处理在>2、0.5~2、0.25~0.5 mm和 < 0.25 mm 4个粒级下分别提高16.99%、36.23%、25.55% 和50.00%,且在0.5~2、0.25~0.5 mm和 < 0.25 mm 3个粒级下差异显著(P < 0.05);V-C处理在4个粒级下分别提高15.69%、21.01%、17.52% 和33.61%,且在 < 0.25 mm粒级下差异显著(P < 0.05);P-C处理在4个粒级下分别提高11.11%、21.74%、20.44%和33.61%,且在0.25~0.5 mm和 < 0.25 mm两个粒级下差异显著(P < 0.05)。不同团聚体粒级之间GRSP的含量则无明显规律。从EEGRSP和T-GRSP综合来看,F-F处理GRSP含量最高,F-C处理最低。
|
|
表 3 不同种植模式下各粒级团聚体中GRSP和SOC含量(mg·g-1) Table 3 Contents of GRSP and SOC in aggregates of each size under different planting patterns(mg·g-1) |
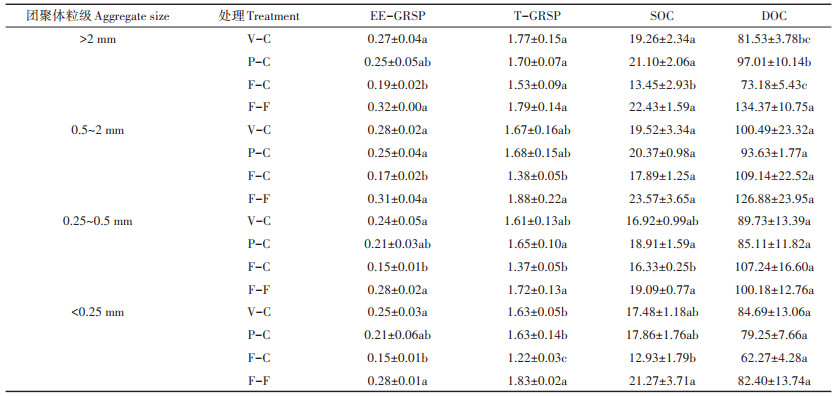
不同处理土壤中SOC的含量为12.93~23.57 mg· g-1,并且呈现出一定的规律性,即同一粒级不同处理SOC含量由大到小依次为F-F、P-C、V-C、F-C。在>2、0.25~0.5 mm和 < 0.25 mm 3个粒级下,与F-C相比,F-F分别显著提高66.76%、16.90% 和64.50%(P < 0.05),P-C在>2 mm和0.25~0.5 mm两个粒级下显著高于F-C(P < 0.05),V-C只在>2 mm粒级下显著高于F-C(P < 0.05)。同一处理不同粒级间SOC含量没有明显规律。不同处理土壤中DOC的含量为62.27~ 134.37 mg·kg-1,F-F处理只在部分粒级高于其他处理。在>2 mm粒级下,F-F显著高于其他3种处理(P < 0.05),P-C显著高于F-C(P < 0.05);在其他3个粒级下各处理间则没有显著差异。从不同粒级团聚体中DOC的含量来看,< 0.25 mm粒级中DOC含量最少,说明本试验土壤中≥0.25 mm粒级团聚体更利于DOC的保存。
2.3 GRSP、SOC和团聚体的结构方程模型分析为明确GRSP对SOC和团聚体稳定性的影响,通过建立结构方程模型(图 1)对相关指标进行分析,χ2/ df、P值、RMSEA、CFI、TLI和GFI表明,该模型适配良好,能代表自变量和因变量的关系。该模型解释了T-GRSP对SOC和DOC都有直接的正面影响,路径系数分别为0.76和0.50;SOC对>0.25 mm水稳性团聚体(WSA>0.25)和MWD有直接正面影响,路径系数分别为0.23和0.15;EE - GRSP对T - GRSP的路径系数为0.90、DOC对WSA>0.25的路径系数为0.83,WSA>0.25对MWD的路径系数为0.96,这些自变量对因变量均有直接的正面影响。从土壤各因子对MWD的直接效应和间接效应看(表 4),各因子对MWD的总效应由大到小依次为WSA>0.25>DOC>T - GRSP>SOC>EE - GRSP。

|
橙色箭头表示正面影响,蓝色箭头表示负面影响,箭头旁边的数字表示路径系数 The orange arrow represents the positive impact, the blue arrow represents the negative impact, and the number next to the arrow represents the path coefficient 图 1 GRSP、SOC和团聚体的结构方程模型 Figure 1 Structural equation model of GRSP, SOC and aggregate |
|
|
表 4 土壤各因子对MWD的直接和间接效应 Table 4 Direct and indirect effects of various soil factors on MWD |

线性回归表明(图 2和图 3),不同团聚体粒级下EE-GRSP与SOC呈正相关关系,但不同粒级下的相关系数大小有所差异,>2、0.5~2 mm和 < 0.25 mm 3个粒级下的相关系数较高,分别为0.69、0.62和0.73,达显著水平,在0.25~0.5 mm中相关系数则较低。不同团聚体粒级下T-GRSP与SOC均呈正相关关系,但不同粒级下的相关系数大小有所差异,在0.5~2 mm和 < 0.25 mm两个粒级下的相关系数较高,分别为0.84和0.76,达显著水平,在>2 mm和0.25~0.5 mm两个粒级下相关系数则较低。从GRSP和SOC数值的分布情况看,在0.5~2 mm粒级下GRSP和SOC的含量比在其他粒级下高。

|
*表示P < 0.05,**表示P < 0.01,NS表示差异不显著,n=12,下同 * means P < 0.05, ** means P < 0.01, NS means the difference is not significant, n=12. The same below 图 2 不同团聚体粒级下EE-GRSP与SOC的关系 Figure 2 The relationship between EE-GRSP and SOC at different aggregate size |

|
图 3 不同团聚体粒级下T-GRSP与SOC的关系 Figure 3 The relationship between T-GRSP and SOC at different aggregate size |
土壤团聚体是土壤结构的基本组成单位[20],是土壤养分存在的主要场所,团聚体结构稳定的土壤可以大幅提高土壤水的渗透性、土壤稳定性并能提高防止自然侵蚀的能力。同时,土壤团聚体合理的多孔结构也为作物的生长提供了必要的空间和良好的通风通道。土壤团聚体也是SOC的重要载体和维持土壤有机质稳定性的物质基础[21],稳定的团聚体可以为其中存在的有机物提供有效的保护。休耕是提高团聚体稳定性行之有效的方法[10, 22-23],与种植作物相比,休耕能提高SOC和全氮的含量[24],同时,轮作也可以提高SOC含量、改善土壤物理性质、提高作物生产力[25]。本试验测定了不同种植模式下水稳性团聚体组成特征,以及团聚体稳定性指标MWD,与其他3种种植模式相比,全年休耕能显著增加土壤中大团聚体(>0.25 mm)含量,提高团聚体稳定性。该结果与前人研究规律一致[6, 15],其原因在于,种植玉米时的机械耕作会增加耕作层土壤的扰动,破坏团聚体结构,进而影响团聚体稳定性的恢复,因此,短期休耕能在一定程度上提高土壤团聚体稳定性,并减少土壤养分的流失,从而恢复由于长期高强度耕作而损失的地力。
丛枝菌根真菌与其他土壤微生物可起到对土壤碳周转的作用[26]。不同土壤耕作方式对土壤条件的改变会对土壤特性产生复杂影响,从而改变土壤环境、影响土壤中微生物多样性和活性,最终对土壤中GRSP含量产生影响[27]。相较于种植大田作物,短期休耕能够提高土壤中T-GRSP的含量[9],作物轮作和残留物管理能显著提高农耕地土壤GRSP含量[6]。本试验测定了不同种植模式下团聚体粒级中的GRSP及SOC的含量,相比F-C,P-C和F-F能在一定程度上提高GRSP和SOC的含量,研究结果与前人一致[28-29]。休耕是实现农业可持续利用的重要措施,其不仅能改善土壤质量,而且有助于土壤中SOC更好地储存[30],一方面,因为休耕减少土壤的翻动,降低了SOC的矿化;另一方面,作物生长需要各种养分,除了从人工施肥中获取,也会从土壤中吸收一部分养分,从而导致种植作物的土壤养分含量降低,而休耕地中的土壤养分则不会被作物所吸收,所以休耕相比轮作能提高土壤养分含量。绿肥-轮作模式中,绿肥成熟后将之翻压到地里,其根系及有机体能够增加土壤微生物多样性[31],进而改善土壤质量,有利于土壤固碳和GRSP的累积。因此,合理的轮作休耕模式能够提高土壤有机质,提升土地生产力,从而达到更好的生态效益和经济效益。
肖玖军等[32]基于结构方程模型对SOC、GRSP和团聚体稳定性指标进行建模分析,发现EE-GRSP、T-GRSP和SOC对团聚体稳定性均有积极的影响,EE-GRSP是主要驱动因子,T-GRSP是团聚体稳定性的主要影响因子。本研究通过构建EE-GRSP、TGRSP、DOC、SOC和MWD的结构方程模型,解释了T-GRSP对SOC和DOC都有直接的正面影响,而SOC对WSA>0.25和MWD有直接正面影响,尽管该模型能解释一些自变量对因变量的作用,但某些路径系数值并不理想,故未来还需建立更合适的模型继续探索GRSP、SOC、DOC与团聚体稳定性的关系。线性回归进一步表明,GRSP和SOC之间存在高度的正相关性,但不同粒级下的相关系数大小有所差异。该结果与FRANZLUEBBERS等[33]、HE等[34]的结果一致。其原因在于:GRSP是有机碳含量的一部分[35],所以能够参与有机碳的封存,提高有机碳含量;另外,可能是因为GRSP和SOC参与相似的生态过程而形成的高度相关[36],如两者都在土壤团聚体形成中起到重要的胶结作用。
综上所述,团聚体稳定性是评价土壤抗蚀性的指标之一,稳定的团聚体能够保护碳库,减少由于土壤侵蚀造成的SOC流失,SOC又进一步参与形成团聚体,增强土壤团聚体稳定性[37],两者相辅相成。GRSP不仅有利于SOC在土壤生态系统中的积累和循环,还能够促进团聚体的形成。合理的轮作和休耕模式能够提高坡耕地GRSP和SOC含量,通过其胶结作用促进土壤团聚作用,从而提高坡耕地抗侵蚀能力。土壤团聚体进一步提高养分储存能力和微生物多样性,最终能够提高土壤肥力并改善土壤健康状况,这对于土壤侵蚀、气候变化和农业增产都具有积极意义。
4 结论(1)相比其他处理,全年休闲能显著增加大团聚体(>0.25 mm)含量,且全年休闲的MWD值显著高于苕子-玉米轮作,说明休闲能提高土壤团聚体的稳定性。
(2)与休闲-玉米相比,豌豆-玉米轮作、苕子-玉米轮作和全年休闲均能有效增加EE-GRSP和TGRSP的储量,且全年休闲效果更佳。不同处理SOC含量由大到小依次为全年休闲、豌豆-玉米轮作、苕子-玉米轮作、休闲-玉米,且全年休闲的SOC显著高于休闲-玉米种植模式(P < 0.05)。从不同处理看DOC的含量,全年休闲只在部分粒级下高于其他种植模式;从不同粒级团聚体中DOC看,≥0.25 mm大团聚体有利于DOC的储存。
(3)通过构建结构方程模型解释了GRSP对SOC和DOC都有直接的正面影响,且对MWD有间接影响,各因子对MWD的总效应由大到小依次为WSA>0.25> DOC>T-GRSP>SOC>EE-GRSP。相关性分析表明,不同团聚体粒级下GRSP与SOC呈线性正相关关系,但不同粒级下的相关系数大小有所差异。
| [1] |
陈正发, 史东梅, 金慧芳, 等. 基于土壤管理评估框架的云南坡耕地耕层土壤质量评价[J]. 农业工程学报, 2019, 35(3): 256-267. CHEN Z F, SHI D M, JIN H F, et al. Evaluation on cultivated-layer soil quality of sloping farmland in Yunnan based on soil management assessment framework(SMAF)[J]. Transactions of the Chinese Society of Agricultural Engineering, 2019, 35(3): 256-267. |
| [2] |
SIX J, PAUSTIAN K. Aggregate-associated soil organic matter as an ecosystem property and a measurement tool[J]. Soil Biology and Biochemistry, 2014, 68: A4-A9. DOI:10.1016/j.soilbio.2013.06.014 |
| [3] |
FRANZLUEBBERS A J. Water infiltration and soil structure related to organic matter and its stratification with depth[J]. Soil and Tillage Research, 2002, 66(2): 197-205. DOI:10.1016/S0167-1987(02)00027-2 |
| [4] |
ŠARAPATKA B, ALVARADO-SOLANO D P, ČIZMAR D. Can glomalin content be used as an indicator for erosion damage to soil and related changes in organic matter characteristics and nutrients?[J]. Catena, 2019, 181: 104078. DOI:10.1016/j.catena.2019.104078 |
| [5] |
权常欣, 马玲玲, 林钊凯, 等. 广东省森林球囊霉素相关土壤蛋白含量及影响因素[J]. 生态环境学报, 2020, 29(2): 240-249. QUAN C X, MA L L, LIN Z K, et al. Patterns and influence factors of glomalinrelated soil protein in Guangdong forests[J]. Ecology and Environmental Sciences, 2020, 29(2): 240-249. |
| [6] |
SINGH G, BHATTACHARYYA R, DAS T K, et al. Crop rotation and residue management effects on soil enzyme activities, glomalin and aggregate stability under zero tillage in the Indo-Gangetic Plains[J]. Soil and Tillage Research, 2018, 184: 291-300. DOI:10.1016/j.still.2018.08.006 |
| [7] |
SPOHN M, GIANI L. Water-stable aggregates, glomalin-related soil protein, and carbohydrates in a chronosequence of sandy hydromorphic soils[J]. Soil Biology and Biochemistry, 2010, 42(9): 1505-1511. DOI:10.1016/j.soilbio.2010.05.015 |
| [8] |
WU Q S, HE X H, ZOU Y N, et al. Spatial distribution of glomalin-related soil protein and its relationships with root mycorrhization, soil aggregates, carbohydrates, activity of protease and β- glucosidase in the rhizosphere of Citrus unshiu[J]. Soil Biology and Biochemistry, 2012, 45: 181-183. DOI:10.1016/j.soilbio.2011.10.002 |
| [9] |
FOKOM R, ADAMOU S, TEUGWA M C, et al. Glomalin related soil protein, carbon, nitrogen and soil aggregate stability as affected by land use variation in the humid forest zone of south Cameroon[J]. Soil and Tillage Research, 2012, 120: 69-75. DOI:10.1016/j.still.2011.11.004 |
| [10] |
LIU H, WANG X, LIANG C, et al. Glomalin-related soil protein affects soil aggregation and recovery of soil nutrient following natural revegetation on the Loess Plateau[J]. Geoderma, 2020, 357: 113921. DOI:10.1016/j.geoderma.2019.113921 |
| [11] |
CARRIZO M E, ALESSO C A, COSENTINO D, et al. Aggregation agents and structural stability in soils with different texture and organic carbon contents[J]. Scientia Agricola, 2015, 72(1): 75-82. DOI:10.1590/0103-9016-2014-0026 |
| [12] |
SINGH A K, RAI A, SINGH N. Effect of long term land use systems on fractions of glomalin and soil organic carbon in the Indo-Gangetic plain[J]. Geoderma, 2016, 277: 41-50. DOI:10.1016/j.geoderma.2016.05.004 |
| [13] |
OGLE S M, BREIDT F J, PAUSTIAN K. Agricultural management impacts on soil organic carbon storage under moist and dry climatic conditions of temperate and tropical regions[J]. Biogeochemistry, 2005, 72(1): 87-121. DOI:10.1007/s10533-004-0360-2 |
| [14] |
李善家, 王军强, 施志国, 等. 不同基肥处理对玉米土壤酶活性和球囊霉素相关土壤蛋白的影响[J]. 应用与环境生物学报, 2017, 23(2): 357-363. LI S J, WANG J Q, SHI Z G, et al. Effect of different base fertilizer treatments on maize soil enzyme activity and glomalinrelated protein[J]. Chinese Journal of Applied and Environmental Biology, 2017, 23(2): 357-363. |
| [15] |
SEKARAN U, SAGAR K L, KUMAR S. Soil aggregates, aggregate-associated carbon and nitrogen, and water retention as influenced by short and long-term no-till systems[J]. Soil and Tillage Research, 2021, 208: 104885. DOI:10.1016/j.still.2020.104885 |
| [16] |
赵德强, 元晋川, 侯玉婷, 等. 玉米||大豆间作对AMF时空变化的影响[J]. 中国生态农业学报(中英文), 2020, 28(5): 631-642. ZHAO D Q, YUAN J C, HOU Y T, et al. Tempo-spatial dynamics of AMF under maize soybean intercropping[J]. Chinese Journal of EcoAgriculture, 2020, 28(5): 631-642. |
| [17] |
ELLIOTT E T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils[J]. Soil Science Society of America Journal, 1986, 50(3): 627-633. DOI:10.2136/sssaj1986.03615995005000030017x |
| [18] |
WRIGHT S F, UPADHYAYA A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi[J]. Soil Science, 1996, 161(9): 575-586. DOI:10.1097/00010694-199609000-00003 |
| [19] |
王展, 张玉龙, 张良, 等. 冻融次数和含水量对棕壤总有机碳和可溶性有机碳的影响[J]. 农业环境科学学报, 2012, 31(10): 1972-1975. WANG Z, ZHANG Y L, ZHANG L, et al. Effect of freeze/thawing frequency and soil water content on total organic carbon and dissolved organic carbon of burozem[J]. Journal of Agro-Environment Science, 2012, 31(10): 1972-1975. |
| [20] |
王润珑, 徐应明, 李然, 等. 膨润土对镉污染土壤团聚体结构特征及有机碳含量的影响[J]. 农业环境科学学报, 2018, 37(12): 2701-2710. WANG R L, XU Y M, LI R, et al. Effects of bentonite on the characteristics of aggregate structure and organic carbon content in Cd-contaminated soils[J]. Journal of Agro - Environment Science, 2018, 37(12): 2701-2710. DOI:10.11654/jaes.2018-0104 |
| [21] |
ZHAO X, WANG C L, ZHAO J, et al. Glomalin-related soil protein and its relationship with organic carbon and nitrogen in water-stable aggregates in abandoned agricultural lands[J]. Journal of Energy and Natural Resources, 2019, 8(1): 37-44. DOI:10.11648/j.jenr.20190801.16 |
| [22] |
ANDERS M M, BRYE K R, OLK D C, et al. Rice rotation and tillage effects on soil aggregation and aggregate carbon and nitrogen dynamics[J]. Soil Science Society of America Journal, 2012, 76(3): 994-1004. DOI:10.2136/sssaj2010.0436 |
| [23] |
GARCIA R A, LI Y, ROSOLEM C A. Soil organic matter and physical attributes affected by crop rotation under no-till[J]. Soil Science Society of America Journal, 2013, 77(5): 1724-1731. DOI:10.2136/sssaj2012.0310 |
| [24] |
姜小凤, 郭凤霞, 陈垣, 等. 休耕和种植作物对黑麻土壤肥力的影响[J]. 水土保持学报, 2021, 35(1): 229-235. JIANG X F, GUO F X, CHEN Y, et al. Effect of fallowing and planting crops on soil fertility of black flax soil[J]. Journal of Soil and Water Conservation, 2021, 35(1): 229-235. |
| [25] |
黄国勤, 赵其国. 中国典型地区轮作休耕模式与发展策略[J]. 土壤学报, 2018, 55(2): 283-292. HUANG G Q, ZHAO Q G. Mode of rotation/fallow management in typical areas of China and its development strategy[J]. Acta Pedologica Sinica, 2018, 55(2): 283-292. |
| [26] |
PARIHAR M, RAKSHIT A, MEENA V S, et al. The potential of arbuscular mycorrhizal fungi in C cycling: A review[J]. Archives of Microbiology, 2020, 202(7): 1581-1596. DOI:10.1007/s00203-020-01915-x |
| [27] |
王建, 周紫燕, 凌婉婷. 球囊霉素相关土壤蛋白的分布及环境功能研究进展[J]. 应用生态学报, 2016, 27(2): 634-642. WANG J, ZHOU Z Y, LING W T. Distribution and environmental function of glomalin-related soil protein: A review[J]. Chinese Journal of Applied Ecology, 2016, 27(2): 634-642. |
| [28] |
白怡婧, 刘彦伶, 李渝, 等. 长期不同轮作模式对黄壤团聚体组成及有机碳的影响[J]. 土壤, 2021, 53(1): 161-167. BAI Y J, LIU Y L, LI Y, et al. Effects of different long-term rotation patterns on aggregate composition and organic carbon in yellow soil[J]. Soils, 2021, 53(1): 161-167. |
| [29] |
MOITINHO M R, FERNANDES C, TRUBER P V, et al. Arbuscular mycorrhizal fungi and soil aggregation in a no-tillage system with crop rotation[J]. Journal of Plant Nutrition and Soil Science, 2020, 183(4): 482-491. DOI:10.1002/jpln.201900456 |
| [30] |
ROLDAN A, SALINAS-GARCIA J R, ALGUACIL M M, et al. Soil sustainability indicators following conservation tillage practices under subtropical maize and bean crops[J]. Soil and Tillage Research, 2007, 93(2): 273-282. DOI:10.1016/j.still.2006.05.001 |
| [31] |
唐治喜, 高菊生, 宋阿琳, 等. 用宏基因组学方法研究绿肥对水稻根际微生物磷循环功能基因的影响[J]. 植物营养与肥料学报, 2020, 26(9): 1578-1590. TANG Z X, GAO J S, SONG A L, et al. Impact of green manure on microbial phosphorus cycling genes in rice rhizosphere as investigated by metagenomics[J]. Journal of Plant Nutrition and Fertilizers, 2020, 26(9): 1578-1590. |
| [32] |
肖玖军, 邢丹, 毛明明, 等. AM真菌对桑树根围土壤团聚体的影响机制[J]. 土壤学报, 2020, 57(3): 773-782. XIAO J J, XING D, MAO M M, et al. Mechanism of arbuscular mycorrhizal fungal affecting soil aggregates in rhizosphere of mulberry(Morus alba)[J]. Acta Pedologica Sinica, 2020, 57(3): 773-782. |
| [33] |
FRANZLUEBBERS A J, WRIGHT S F, STUEDEMANN J A. Soil aggregation and glomalin under pastures in the southern Piedmont USA[J]. Soil Science Society of America Journal, 2000, 64(3): 1018-1026. DOI:10.2136/sssaj2000.6431018x |
| [34] |
HE J D, CHI G G, ZOU Y N, et al. Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange[J]. Applied Soil Ecology, 2020, 154: 103592. DOI:10.1016/j.apsoil.2020.103592 |
| [35] |
杨振亚, 阙弘, 朱雪竹, 等. 几种丛枝菌根真菌对菲污染土壤中球囊霉素含量的影响[J]. 农业环境科学学报, 2016, 35(7): 1338-1343. YANG Z Y, QUE H, ZHU X Z, et al. Impacts of arbuscular mycorrhizal fungi on glomalin content in soils contaminated with phenanthrene[J]. Journal of Agro-Environment Science, 2016, 35(7): 1338-1343. |
| [36] |
WILSON G W T, RICE C W, RILLIG M C, et al. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments[J]. Ecology Letters, 2009, 12(5): 452-461. DOI:10.1111/j.1461-0248.2009.01303.x |
| [37] |
NAUTIYAL P, RAJPUT R, PANDEY D, et al. Role of glomalin in soil carbon storage and its variation across land uses in temperate Himalayan regime[J]. Biocatalysis and Agricultural Biotechnology, 2019, 21: 101311. DOI:10.1016/j.bcab.2019.101311 |