 2018, Vol. 37
2018, Vol. 37



2. 山西省农业科学院谷子研究所, 山西 长治 046000
2. Institute of Millet Research, Shanxi Academy of Agricultural Science, Changzhi 046000, China
谷子是我国北方地区的主栽作物之一,在农业生产中占据重要地位,但由于耕地有限、种植模式不合理的影响,导致连作障碍普遍发生。连作障碍是指在同一块土壤中连续种植同种作物,导致作物长势变弱、产量降低、品质下降等现象[1]。研究表明,发生连作障碍的原因主要有以下几点:土壤养分失衡;土壤pH值降低;土壤微生物群落结构变化,有害微生物增加[2-3]。轮作是农业生产中常用的一种种植制度,合理轮作能够平衡土壤养分,改变土壤理化性质,增加作物产量,有效避免或减少连作障碍的发生。
土壤酶参与土壤养分的固定和释放,以及各种氧化还原反应,其活性是评价土壤肥力的重要参数之一[4]。土壤微生物主导土壤生态系统中的养分循环和能量流动,对于维持系统稳定性和可持续性有重要意义[5]。研究表明,轮作不仅有助于提高土壤酶活性,对土壤微生物组成和结构多样性也有明显促进作用。李锐等[6]发现长期棉花连作使农田土壤酶活性下降,微生物多样性降低,而轮作可提高土壤过氧化氢酶(CAT)、多酚氧化酶(PPO)和蔗糖酶活性以及微生物群落结构多样性;于高波等[7]报道小麦、毛苕子与黄瓜轮作明显改善了根际细菌组成,且土壤CAT、PPO和脲酶活性显著升高;王劲松等[8]研究表明高粱-苜蓿轮作能增加土壤蔗糖酶活性和微生物多样性。
迄今为止,关于轮作能改善土壤质量、缓解连作障碍的报道很多[6-9],但谷子作为山西省优势特色杂粮作物,具有耐瘠薄、适应性强等特点,是否可以建立某种轮作模式避免连年种植带来的不利影响,尚待研究。因此,本研究设置谷子连作、马铃薯-玉米-谷子轮作、玉米-大豆-谷子轮作和大豆-马铃薯-谷子轮作4个处理,监测不同种植模式下土壤养分和酶活性变化特征,并利用Illumina MiSeq测序技术对谷子根际土壤细菌多样性进行分析,明确不同种植模式对土壤生态环境的影响,为谷田种植模式优化和农业可持续生产提供依据。
1 材料与方法 1.1 试验概况试验在山西省长治市山西省农业科学院谷子研究所的谷子试验田进行(113°06′E,36°11′N),试验地所在区域日平均气温28 ℃,夜平均气温18 ℃。供试土壤为浅黏绵垆土,含碱解氮27.56 mg·kg-1、有效磷15.01 mg·kg-1、速效钾93.30 mg·kg-1,有机质15.43 g·kg-1,pH 7.14。
试验开展前该田块为谷子连作土壤,于2012年开始轮作试验,选用的4种轮作作物分别是:谷子(Setaria italica,Si)、玉米(Zea mays,Zm)、马铃薯(Solanum tuberosum,St)和大豆(Glycine max,Gm)。共设四种不同种植模式:谷子连作(Si-Si-Si)、马铃薯-玉米-谷子轮作(St-Zm-Si)、玉米-大豆-谷子轮作(Zm-Gm-Si)和大豆-马铃薯-谷子轮作(Gm-St-Si),具体试验方案见表 1。每种种植模式3次重复,随机区组排列,每个小区面积为30 m2,株间距35 cm,每公顷留苗30万株。2012—2015年各小区管理施肥相同,施肥水平均为氮肥(N)150 kg·hm-2、磷肥(P2O5)150 kg·hm-2、钾肥(K2O)30 kg·hm-2,播种前将40%氮肥和全部磷肥、钾肥作为基肥施入,其余氮肥于谷子拔节期追肥,谷子追肥时给同期田间种植的马铃薯、玉米和大豆施用等量氮肥。
|
|
表 1 试验设计 Table 1 Design of experiment |

于2014年谷子拔节期,每个小区随机选3个采样点,每个样点随机选取长势一致的5棵植株,挖取植株根周5 cm范围内的全部根系,用手轻抖植株根系,去除附着土,用软毛刷刷取根部的根际土,过20目筛后装于自封袋,-80 ℃保存,用作土壤微生物多样性检测。同期,随机选各小区3个点,采集0~20 cm的耕层土壤,去除杂物,过20目筛后装于自封袋,4 ℃保存,用作土壤基本理化性质和酶活性测定。
1.2.2 土壤pH和养分测定土壤pH测定采用电位法。土壤养分测定参照鲁如坤[10]的方法,采用碱解扩散法测定土壤碱解氮含量,钼锑抗比色法测定土壤有效磷含量,火焰光度法测定土壤速效钾含量,重铬酸钾法测定土壤有机质。
1.2.3 土壤酶活性测定土壤酶活性检测参照关松荫[11]的方法。过氧化氢酶(CAT)活性采用高锰酸钾滴定法测定,以20 min后1 g土壤消耗1 mL 0.02 mol·L-1高锰酸钾所需要的酶量定义为1个酶活力单位(mg·g-1,FW);多酚氧化酶(PPO)活性采用邻苯三酚法测定,以2 h后1 g土壤中产生1 mg紫色没食子素所需要的酶量定义为1个酶活力单位(mg·g-1,FW);纤维素酶活性采用硝基水杨酸比色法测定,以72 h、1 g土壤中产生1 mg葡萄糖所需要的酶量定义为1个酶活力单位(mg·g-1,FW);蔗糖酶活性采用3,5-二硝基水杨酸法测定,以24 h、1 g土壤中产生1 mg葡萄糖所需要的酶量定义为1个酶活力单位(mg·g-1,FW)。
1.2.4 产量测定2014年谷子成熟时按小区实收记产,然后折算成公顷产量。
1.2.5 土壤微生物多样性检测使用OMEGA公司生产的E.Z.N.A Soil DNA试剂盒,按照试剂盒的使用说明提取和纯化土壤微生物基因组DNA。将纯化后的基因组DNA作为PCR模板,扩增细菌16S rDNA的V4区,使用的引物是520 F:5′-AYTGGGYDTAAAGNG-3′;802R:5′- TACNVGGGTATCTAATCC-3′。PCR扩增条件为:95 ℃预变性4 min,94 ℃变性30 s,55 ℃退火45 s,72 ℃延伸30 s,25个循环后,72 ℃延伸5 min。琼脂糖凝胶电泳检测PCR产物,目的条带用Qiagen公司生产的QIAquick Gel Extraction Kit凝胶回收试剂盒切胶回收。回收的DNA送上海派森诺生物科技股份有限公司进行测序,测序平台为Illumina MiSeq。对原始数据进行优化,去除引物序列、低于150 bp的序列和低质量的序列;只保留重叠超过10 bp且没有错配的序列。在0.97相似度下利用QIIME软件对操作分类单元(Operational Taxonomic Units,OTU)进行聚类分析,并制作维恩图;基于样品的OTU数量进行Alpha多样性分析,计算Chao、Ace、Shannon指数[12-14];在门和属水平上对样品OTU进行聚类,对聚类后各样品所含的不同OTU序列做柱状图和热图。
1.3 数据处理试验数据采用Excel 2010进行数据统计,采用SPSS 19.0进行数据分析,通过Duncan方法对不同处理组之间的数据进行多重比较。
2 结果与分析 2.1 土壤pH和土壤养分在三种轮作模式下,土壤pH值和碱解氮、有效磷、速效钾、有机质的含量均发生改变(图 1)。与连作相比,pH值在三个轮作处理中均升高(图 1A);碱解氮含量Gm-St-Si显著升高,St-Zm-Si、Zm-Gm-Si无显著改变(图 1B);有效磷含量Gm-St-Si中最高,Zm-Gm-Si差异不显著,St-Zm-Si降低(图 1C);速效钾含量在Gm-St-Si和Zm-Gm-Si中均升高,其中Gm-St-Si最高,增幅34.4%,St-Zm-Si差异不显著(图 1D);有机质含量三个轮作处理均显著升高(图 1E)。

|
不同字母表示差异显著(P<0.05)。下同 Different letters indicate the significant differences(P < 0.05). The same below 图 1 四种种植模式下的土壤pH(A)和碱解氮(B)、有效磷(C)、速效钾(D)、有机质(E)含量 Figure 1 Soil pH (A) and the contents of alkaline hydrolysis nitrogen(B), available phosphorus (C), potassium (D), and organic matter (E) under four cropping systems |
酶活性检测结果显示,轮作会影响土壤CAT、PPO、纤维素酶和蔗糖酶活性。CAT活性在轮作处理中显著提高,但三个轮作处理间未表现显著差异(图 2A);PPO活性在St-Zm-Si和Gm-St-Si中最高,Zm-Gm-Si次之,连作最低(图 2B);纤维素酶活性呈现Gm-St-Si>Zm-Gm-Si>St-Zm-Si>Si-Si-Si的趋势,其中Gm-St-Si是连作的3.25倍(图 2C);蔗糖酶活性在Gm-St-Si、Zm-Gm-Si和St-Zm-Si中显著升高,分别比连作高29.48%、13.23%和8.91%(图 2D)。综上表明,与谷子连作相比,轮作后各土壤酶活性均处于较高水平,其中Gm-St-Si轮作的效果最显著。

|
图 2 四种种植模式下的土壤酶活性 Figure 2 Soil enzyme activities under four cropping systems |
统计四种种植模式下谷子的产量,结果显示,轮作处理的谷子产量均高于连作,但只有Gm-St-Si达到显著水平,较连作高37.11%,Zm-Gm-Si和St-Zm-Si比连作高31.08%和27.52%(图 3)。

|
图 3 2014年谷子产量 Figure 3 Yield of millet in 2014 |
在相似度0.97的水平上对测序所得序列进行聚类,共得到8750个OTU。如图 4所示,Si-Si-Si、StZm-Si、Zm-Gm-Si和Gm-St-Si组的OTU数分别为4519、4692、4970个和5745个,具体排序为Gm-St-Si>Zm-Gm-Si>St-Zm-Si>Si-Si-Si,三个轮作处理的OTU数均高于连作,Gm-St-Si处理最高,为连作的1.27倍。

|
图 4 四种种植模式下土壤OTU分布维恩图 Figure 4 Venn diagram showing the distribution of OTU of soil samples under four cropping systems |
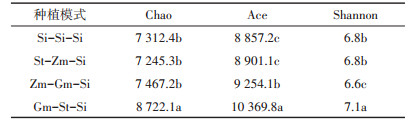
本研究考察的Alpha多样性指标包括丰富度指数(Chao、Ace)和多样性指数(Shannon)(表 2)。Chao值在Gm-St-Si中最高,St-Zm-Si、Zm-Gm-Si与连作间无显著差异。与连作相比,Ace值在Gm-St-Si和Zm-Gm-Si中显著增加,St-Zm-Si差异不显著;Shannon值在Gm-St-Si中显著升高,Zm-Gm-Si中显著下降,St-Zm-Si差异不显著。与谷子连作比较,Gm-St-Si的Chao值、Ace值和Shannon值均呈现升高趋势。
|
|
表 2 四种种植模式下根际土壤细菌群落的Alpha多样性指数 Table 2 Alpha-diversity index of rhizosphere bacterial community under four cropping systems |
检出的细菌主要包括8个门,Verrucomicrobia(疣微菌门)、Actinobacteria(放线菌门)、Acidobacteria(酸杆菌门)、Proteobacteria(变形菌门)、Bacteroidetes(拟杆菌门)、Planctomycetes(浮霉菌门)、Gemmatimonadetes(芽单胞菌门)和Chloroflexi(绿弯菌门),四种种植模式下根际土壤细菌各门类的丰度有明显差异(图 5)。与连作相比,三个轮作处理含有更丰富的疣微菌门、酸杆菌门和浮霉菌门。其中,疣微菌门在Zm-Gm-Si和Gm-St-Si中较丰富,酸杆菌门在Gm-St-Si中含量较多,浮霉菌门在Gm-St-Si中丰度最高。

|
图 5 基于门水平的细菌组成分布图 Figure 5 Bacteria community distribution based on the level of phylum |
在属的水平上对样品所含菌属进行聚类,分析其细菌组成的相似性和多样性。由图 6可知,不同作物轮作后土壤的细菌群落结构发生改变,其中Si-Si-Si和St-Zm-Si聚为一支,都含有丰富的厚壁菌门和放线菌门,Zm-Gm-Si和Gm-St-Si聚为一支,以变形菌门和浮霉菌门为主。与连作相比,三个轮作处理的Planctomyces、Gemmata、Flavisolibacter、Luteolibacter菌群丰度增加,Gm-St-Si处理增加的幅度最大,其中Planctomyces丰度比连作提高了1.16倍,Gemmata丰度提高了1.14倍,Flavisolibacter丰度是连作的1.87倍,Luteolibacter丰度是连作的3.89倍;而Flavobacterium、Luteimonas菌群丰度在Gm-St-Si中降低(图 7)。

|
图 6 基于属水平的细菌组成聚类分析 Figure 6 Cluster analysis of bacteria community based on the level of genus |

|
在Gm-St-Si处理中,Luteimonas、Flavobacterium菌群丰度降低,Planctomyces、Gemmata、Flavisolibacter、Luteolibacter菌群丰度增加 图 7 基于属水平的细菌相对丰度 Figure 7 Relative abundance of the dominant genus |
适当的轮作方式能改善土壤理化性状,有效避免土壤酸化,还能均衡土壤中的营养元素,平衡养分含量[1-2]。本研究结果表明,轮作后土壤有机质含量和pH值均有所升高,土壤肥力增加,土壤微生物多样性提高,有利于作物的生长发育和增产。在种植大豆的轮作组中,土壤碱解氮含量显著增加,可能与豆类作物的固氮作用有关,大豆能固定空气中的无机氮,增加土壤氮含量[15];而在St-Zm-Si处理中,土壤有效磷含量较低,可能是前茬种植玉米对磷肥消耗较多所致。前人研究报道,前茬种植玉米会导致农田磷含量下降[16],我们的结果与此一致。
土壤理化性状和酶活性是衡量土壤肥力的重要指标,两者共同作用推动土壤代谢过程,影响作物生长,土壤酶来源于植物根系分泌物和土壤微生物,易受土壤环境因子的影响[4]。轮作为农田生态系统提供了较高的植物多样性,使土壤微生物多样性增加,并提高土壤酶活性。前人研究表明,土壤中CAT、PPO活性与植株抗病性呈正相关;纤维素酶和蔗糖酶是碳循环中的关键酶,对土壤有机质利用有重要意义[6]。本研究结果显示,轮作使土壤CAT、PPO、纤维素酶和蔗糖酶活性升高,有利于增强植株抗病性和对土壤养分的利用。
土壤微生物参与土壤多种重要的生物化学过程,其群落的结构及组成直接影响土壤微生态系统稳定和平衡,进而影响作物正常生长发育。土壤微生物主要由细菌、真菌和放线菌组成,其中细菌是数量最多的一个类群,参与有机质分解、氨化反应等多种生物学过程[8]。在土壤中,根际微生物与植物根系分泌物之间的相互作用是一个重要的过程,根系通过分泌多种化合物,对根际微生物发挥促进或抑制作用,进而对微生物群落结构产生影响[5]。轮作作为一种常见的农业技术,可使几种作物的根系分泌物、作物残体在土壤中积累,丰富了土壤微生物的营养物质,促进根际微生物群落的多样性。在本研究中发现三种轮作模式下谷子根际微生物多样性提高,与前人结果一致。
谷子根际土壤细菌多样性分析发现,轮作处理的细菌数目和多样性指数明显高于连作,细菌群落结构组成亦明显不同。轮作处理中疣微菌门、酸杆菌门和浮霉菌门含量明显升高。有研究表明,疣微菌门和酸杆菌门能有效分解有机物、提高植株抗逆能力[17];浮霉菌门在植物营养成分的吸收,特别是碳源、氮源的利用方面具有重要作用[18]。另外,轮作使部分功能菌的数量增加,如Planctomyces、Gemmata、Flavisolibacter、Luteolibacter等。Planctomyces可将多糖分解为小分子糖类被植物利用[19];Gemmata能增加脲酶的活性[20],脲酶是一种酰胺酶,可酶促分解有机物,脲酶活性升高能提高植物对有机物的利用率;Flavisolibacter能促进土壤磷素循环[21];Luteolibacter参与植物对有机质的分解利用过程,对环境中碳循环有重要意义,还可能对某些植物真菌性病害有拮抗作用[22]。此外,Flavobacterium、Luteimonas含量在轮作组中明显减少,而Flavobacterium可能和一些植物病害的发生有关,其丰度与谷子黑穗病的发病率呈正相关[23],且在发病小麦的根际土壤中富集[24]。综上表明,轮作使谷子根际土壤中细菌的群落结构发生变化,有益菌的数量明显增加,有害菌的数量减少,对于促进营养成分吸收、提高有机质利用率、增强植株抗病性有重要意义,也是谷子增产的有利条件。
4 结论几种不同的轮作方式对土壤理化性质、酶活性和根际细菌多样性等均产生影响,其中大豆-马铃薯-谷子轮作后谷子根际土壤细菌多样性指数最高,土壤中富含Planctomyces、Gemmata、Flavisolibacter、Luteolibacter等有益功能菌,具有更高的土壤养分和土壤酶活性,获得较高谷子产量。综上表明,利用大豆-马铃薯-谷子轮作有助于改善谷田生态环境,增强土壤肥力,促进谷子增产。
| [1] |
Qin S, Yeboah S, Li C, et al. Breaking continuous potato cropping with legumes improves soil microbial communities, enzyme activities and tuber yield[J]. PLoS One, 2017, 12(5): e0175934. DOI:10.1371/journal.pone.0175934 |
| [2] |
Asuming-Brempong S, Gantner S, Adiku S G K, et al. Changes in the biodiversity of microbial populations in tropical soils under different fallow treatments[J]. Soil Biology and Biochemistry, 2008, 40(11): 2811-2818. DOI:10.1016/j.soilbio.2008.08.010 |
| [3] |
Xiao X M, Cheng Z H, Meng H W, et al. Intercropping with garlic alleviated continuous cropping obstacle of cucumber in plastic tunnel[J]. Acta Agriculturae Scandinavica, 2012, 62(8): 696-705. |
| [4] |
董艳, 董坤, 郑毅, 等. 种植年限和种植模式对设施土壤微生物区系和酶活性的影响[J]. 农业环境科学学报, 2009, 28(3): 527-532. DONG Yan, DONG Kun, ZHENG Yi, et al. Soil microbial community and enzyme activities in greenhouse with different cultivation years and planting system[J]. Journal of Agro-Environment Science, 2009, 28(3): 527-532. DOI:10.3321/j.issn:1672-2043.2009.03.020 |
| [5] |
吴林坤, 林向民, 林文雄. 根系分泌物介导下植物-土壤-微生物互作关系研究进展及展望[J]. 植物生态学报, 2014, 38(3): 298-310. WU Lin-kun, LIN Xiang-min, LIN Wen-xiong. Advances and perspective in research on plant-soil-microbe interactions mediated by root exudates[J]. Chinese Journal of Plant Ecology, 2014, 38(3): 298-310. |
| [6] |
李锐, 刘瑜, 褚贵新, 等. 不同种植方式对绿洲农田土壤酶活性与微生物多样性的影响[J]. 应用生态学报, 2015, 26(2): 490-496. LI Rui, LIU Yu, CHU Gui-xin, et al. Effects of different cropping patterns on soil enzyme activities and soil microbial community diversity in oasis farmland[J]. Chinese Journal of Applied Ecology, 2015, 26(2): 490-496. |
| [7] |
于高波, 吴凤芝, 周新刚. 小麦、毛苕子与黄瓜轮作对土壤微生态环境及产量的影响[J]. 土壤学报, 2011, 48(1): 175-184. YU Gao-bo, WU Feng-zhi, ZHOU Xin-gang. Effects of rotations of cucumber with wheat and hairy vetch on soil micro-ecological environment and its yield[J]. Acta Pedologica Sinica, 2011, 48(1): 175-184. |
| [8] |
王劲松, 樊芳芳, 郭珺, 等. 不同作物轮作对连作高粱生长及其根际土壤环境的影响[J]. 应用生态学报, 2016, 27(7): 2283-2291. WANG Jin-song, FAN Fang-fang, GUO Jun, et al. Effects of different crop rotations on growth of continuous cropping sorghum and its rhizosphere soil micro-environment[J]. Chinese Journal of Applied Ecology, 2016, 27(7): 2283-2291. |
| [9] |
Tan Y, Cui Y S, Li H Y, et al. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panax notoginseng cropping practices[J]. Microbiological Research, 2017, 194: 10-19. DOI:10.1016/j.micres.2016.09.009 |
| [10] |
鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. LU Ru-kun. Methods of soil agricultural chemical analysis[M]. Beijing: China Agricultural Science and Technology Press, 2000. |
| [11] |
关松荫. 土壤酶及其研究方法[M]. 北京: 农业出版社, 1986: 303-312. GUAN Song-yin. Soil enzyme and its research methods[M]. Beijing: Agricultural Publishing House, 1986: 303-312. |
| [12] |
Chao A. Nonparametric estimation of the number of classes in a population[J]. Scandinavian Journal of Statistics, 1984, 11(4): 265-270. |
| [13] |
Chao A, Lee S M. Estimating the number of classes via sample coverage[J]. Publications of the American Statistical Association, 1992, 87(417): 210-217. DOI:10.1080/01621459.1992.10475194 |
| [14] |
Soetaert K, Heip C. Sample-size dependence of diversity indices and the determination of sufficient sample size in a high-diversity deepsea environment[J]. Marine Ecology Progress, 1990, 59: 305-307. DOI:10.3354/meps059305 |
| [15] |
秦舒浩, 曹莉, 张俊莲, 等. 轮作豆科植物对马铃薯连作田土壤速效养分及理化性质的影响[J]. 作物学报, 2014, 40(8): 1452-1458. QIN Shu-hao, CAO Li, ZHANG Jun-lian, et al. Effect of rotation of leguminous plants on soil available nutrients and physical and chemical properties in continuous cropping potato field[J]. Acta Agronomica Sinica, 2014, 40(8): 1452-1458. |
| [16] |
孙伟红, 劳秀荣, 董玉良, 等. 小麦-玉米轮作体系中秸秆还田对产量及土壤钾素肥力的影响[J]. 作物杂志, 2004, 4: 14-16. SUN Wei-hong, LAO Xiu-rong, DONG Yu-liang, et al. Effects of straw mulching on yield and soil potassium fertility in wheat-maize rotation system[J]. Crops, 2004, 4: 14-16. |
| [17] |
Banerjee S. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil[J]. Soil Biology and Biochemistry, 2016, 97: 188-198. DOI:10.1016/j.soilbio.2016.03.017 |
| [18] |
Bhattacharyya P, Roy K S, Nayak A K, et al. Metagenomic assessment of methane production-oxidation and nitrogen metabolism of long term manured systems in lowland rice paddy[J]. Science of the Total Environment, 2017, 586: 1245-1253. DOI:10.1016/j.scitotenv.2017.02.120 |
| [19] |
Erbllgin O, Mcdonald K L, Kerfeld C A. Characterization of a planctomycetal organelle:A novel bacterial microcompartment for the aerobic degradation of plant saccharides[J]. Applied and Environmental Microbiology, 2014, 80(7): 2193-2205. DOI:10.1128/AEM.03887-13 |
| [20] |
Zhang X X, Zhang R J, Gao J S, et al. Thirty-one years of rice-ricegreen manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria[J]. Soil Biology and Biochemistry, 2017, 104: 208-217. DOI:10.1016/j.soilbio.2016.10.023 |
| [21] |
Hong C, Si Y X, Xing Y, et al. Illumina MiSeq sequencing investigation on the contrasting soil bacterial community structures in different iron mining areas[J]. Environmental Science and Pollution Research, 2015, 22(14): 10788-10799. DOI:10.1007/s11356-015-4186-3 |
| [22] |
Rocha U N, Elsas J D, Overbeek L S. Real-time PCR detection of Holophagae(Acidobacteria)and Verrucomicrobia subdivision 1 groups in bulk and leek(Allium porrum) rhizosphere soils[J]. Journal of Microbiological Methods, 2010, 83(2): 141-148. DOI:10.1016/j.mimet.2010.08.003 |
| [23] |
Han Y S, Xu L X, Liu L Q, et al. Illumina sequencing reveals a rhizosphere bacterial community associated with foxtail millet smut disease suppression[J]. Plant and Soil, 2017, 410(1): 411-421. |
| [24] |
Gardener M S, Weller D M. Changes in populations of rhizosphere bacteria associated with take-all disease of wheat[J]. Applied and Environmental Microbiology, 2001, 67(10): 4414-4425. DOI:10.1128/AEM.67.10.4414-4425.2001 |