 2018, Vol. 37
2018, Vol. 37文章信息
- 魏甲彬, 徐华勤, 周玲红, 成小琳, 唐先亮, 傅志强, 唐启源, 唐剑武
- WEI Jia-bin, XU Hua-qin, ZHOU Ling-hong, CHENG Xiao-lin, TANG Xian-liang, FU Zhi-qiang, TANG Qi-yuan, TANG Jian-wu
- “双季稻-冬闲田”生态系统碳交换动态变化及其影响因素
- Seasonal variation in carbon exchange and its modulating factors of a double cropping rice ecosystem in Southern China
- 农业环境科学学报, 2018, 37(5): 1035-1044
- Journal of Agro-Environment Science, 2018, 37(5): 1035-1044
- http://dx.doi.org/10.11654/jaes.2017-1179
文章历史
- 收稿日期: 2017-08-29
- 录用日期: 2017-12-19






由于大气CO2浓度的升高与全球气候变化之间具有密切的相关性,为明确各类生态系统在大气CO2浓度升高过程中扮演的角色及其固碳减排潜力,各类生态系统与大气圈之间的碳交换研究至关重要[1]。作为全球气候变化和区域可持续发展研究的核心内容之一,陆地生态系统碳循环规律已成为国际环境领域研究的热点和焦点[2]。农田生态系统作为陆地生态系统的重要组成部分,受人为因素强烈影响和干扰,其CO2的生态系统净交换量变化较大,已成为国际碳循环研究的热点问题之一[3-6]。在生态系统碳交换研究方法中,传统的箱式法(暗箱或明箱)存在测定时对下垫面扰动较大、测定不具备连续性以及时间分辨率低等缺陷,在估算生态系统碳交换中存在较大的不确定性。涡度相关(Eddy covariance)技术能够实现对生态系统尺度碳交换的直接测定[7],是近年来国际上普遍认可的方法,并逐渐应用于农田生态系统碳通量观测研究。近10年来,国内外学者针对农田生态系统净碳交换开展了一系列的相关研究,研究对象主要集中在小麦、水稻、玉米、棉花等单个作物在生长季或者某种耕作措施下土壤呼吸、CO2通量特征及其影响因素等方面,这些研究一方面对农田休闲期碳排放关注较少,另一方面,采用的主要是传统的箱式法进行测定,有关年尺度的农田生态系统高通量的碳交换研究则只有少量报道[8-12],对农田生态系统周年碳平衡的研究不足。
截至2014年全国双季稻种植面积达618.18万hm2[13]。双季稻是湖南省目前主要的水稻种植模式。因此,研究这一地区农田生态系统碳交换的特征及其环境影响因子对区域碳平衡估算具有重要的意义。但关于双季稻区稻田生态系统的碳交换研究到目前为止只有少量报道,且研究方法主要是箱式法,鲜见使用涡度相关法进行全年碳交换研究的报道[14-17]。同时,农田生态系统受区域空间分布、气候与土壤环境、耕作制度、管理措施等差异影响,区域间农田生态系统碳平衡的估算与比较存在不确定因素较多,需要更多的案例研究。鉴于此,本研究通过涡度相关技术,对湖南省浏阳市典型双季稻生态系统进行连续一年的通量观测,探讨其季节性碳交换动态特征及其主要环境影响因子,为评估南方双季稻农田生态系统周年碳平衡提供科学参考。
1 材料与方法 1.1 研究区概况试验于2015年全年在湖南省浏阳市沿溪镇(113°84′E,28°30′N)湖南农业大学教学科研基地进行,该基地所处的湘东山区为丘陵地形,属亚热带季风湿润气候,年平均气温17.3 ℃,1月平均气温5.4 ℃,7月平均气温28.7 ℃,年均降水量1 358.6~1 552.5 mm,土壤类型为潴育性水稻土。其有机碳、全氮、全磷、全钾含量分别为29.1、1.58、0.49、14.62 g·kg-1,pH值5.71。
以涡度相关通量塔为中心,南北长100 m的观测区内地势平坦,且均为水稻田,历年种植制度为稻-稻-冬闲,且水稻秸秆均还田。早稻品种为中嘉早17,于2015年4月21日进行机插秧,7月18日收割,收割后水稻秸秆旋耕进土层;晚稻品种为H优518,于2015年7月21日进行机插秧,10月31日收割,收割后水稻秸秆旋耕进土层。试验期间早晚稻均施氮磷钾复合肥750 kg·hm-2(N:P:K=15:15:15),随后的分蘖肥施尿素375 kg·hm-2。水分管理为当地常规管理。
1.2 研究方法观测系统由涡度相关观测系统和常规气象要素观测系统组成。涡度相关观测系统主要包括CO2/H2O分析及三维超声风速仪(EC150,Campbell)和数据采集器(CR1000,Campbell),分别用于快速测定冠层上方的三维风速、超声虚温、水汽和CO2浓度。系统连续自动采集,采样频率为10 Hz,由数据采集器自动存储在原装进口U盘(16 G)内。常规气象要素观测系统为Vantage pro2型自动气象站,用于测定总辐射、气温、和降水量等。同时,用Onset HOBO温度记录仪(U23-001)长期监控土壤温度(0~5 cm)。以上各项测定值采样周期均为30 min的平均值。另外采用箱式法测定生态系统CO2通量,以此通量数据对涡度相关法数据进行质量控制。箱式法测定间隔为水稻生育期每半个月测定1次,冬闲田期间每个月中旬测定1次,采用超便携式温室气体(CH4、CO2、H2O)自动分析仪(Ultraportable Greenhouse Gas Analyzer,美国Los Gatos Research公司,仪器型号为915-0011-1000)与循环采气透明静态箱相结合。静态箱材质为透明有机玻璃,规格为50 cm×50 cm。每个小区布置一个相同尺寸的正方形底座,四边有凹槽,安装时底座边缘嵌入土中3 cm深。测定时,静态箱置于底座上,凹槽内灌水以确保透明静态箱处于密闭状态,每个小区测定5 min,从上午7点开始用暗箱测定生态系统呼吸,测定时,先于早晨气温较低时用黑布罩住静态箱,测定生态系统呼吸排放的CO2通量,随后分别测定上午和下午的生态系统CO2通量。
叶面积指数(Leaf area index,LAI)采用直尺直接测量的方法。在每次用箱式法测定时,于田间确定5个点,每个点随机测量10株水稻每个叶片的最大叶长与最大叶宽,然后用以下公式计算LAI,以50株水稻平均值作为该次测定的水稻LAI:
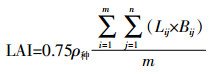 (1)
(1) 式中:n为第j株的总叶片数;m为测定株数;ρ种为种植密度。
1.3 数据处理原始数据取得后,采用eddypro 6.2.0进行30 min通量计算,软件自动进行异常值处理和相关校正。获得的全年通量数据进一步做如下处理:剔除夜间(定义太阳总辐射 < 10 W·m-2为夜间[18])为负值的数据值;剔除明显不符合经验常规的数据值(本文根据箱式法测定的结果剔除>20 μmol·m-2·s-1和 < -30 μmol·m-2·s-1的数据值);剔除降雨时刻的数据值,因为降雨会扰动气体流动甚至导致仪器失灵;剔除摩擦风速 < 0.05 m·s-1对应的通量数据值以消除夜间湍流低对数据质量的影响[19]。
通量观测过程中,受仪器故障、天气状况、大气稳定度和供电系统故障等影响造成大量数据的异常和缺失,数据不可用比例在17%~50%[19-22]。本站点观测时间为2015年1月1日—12月31日,全年有效数据为45%,其中白天有效数据为51%,夜间有效数据为39%。因此,需要进行数据插补。气象数据也有小部分缺失,采用和CO2通量相同的方法策略进行插补。缺失数据插补方法主要包括:平均日变化法、根据特定气象条件查表法、非线性回归法[20]。由于利用全年有效数据建立的线性方程决定系数较小,故本文采用以下方法进行插补:(1) < 2 h缺失数据用线性内插法;(2)>2 h且 < 1 d缺失数据采用平均日变化法,白天取14 d、夜间取7 d的平均时间长[20, 22]。
涡度相关系统观测并通过eddypro计算得到的是30 min的CO2通量数据,这个数据只是陆地生态系统和大气之间的净碳交换量(Net ecosystem exchange,NEE),即总初级生产力(Gross primary productivity,GPP)和生态系统总呼吸(Ecosystem respiration,Reco)之差。但是在冠层尺度的GPP和Reco通常是无法直接观测的,需要通过一定算法将实测NEE进行分解。分解NEE的基本原理是通过夜间观测的NEE(夜间无光合作用,因此净交换量就是生态系统净碳排放,即NEE=Reco)确定Reco温度响应函数的参数,然后将该关系应用于白天确定白天的Reco,进而计算获得白天GPP。本文采用van′t Hoff经验指数模型建立Reco与温度的关系[20]:
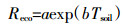 (2)
(2) 式中:a、b为模型建立后的系数,分别表示当温度为0 ℃时的生态系统呼吸和生态系统呼吸对温度的响应参数;Tsoil为5 cm土壤温度;b值与Q10模型具有一致的关系,因此Q10值采用以下模型计算:
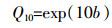 (3)
(3) 利用计算得到的白天Reco值,可以得到生态系统全年GPP的值:
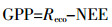 (4)
(4) 最后,采用米氏方程进行光合有效辐射(Photosynthetically active radiation,PAR)与NEE的相关性分析[20]:
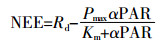 (5)
(5) 式中:Rd为白天的呼吸速率;Pmax为光饱和时的光合速率;Km为拟合常数;α为初始表观量子效率,α=-Pmax /Km;PAR为光合有效辐射,μmol·m-2·s-1。本研究中的PAR由太阳总辐射计算而来[23]。
2 结果与分析 2.1 “双季稻-冬闲田”生态系统气象与环境因子分析图 1显示了2015年试验站点的PAR、降雨量、气温以及5 cm土壤温度的季节性动态变化。PAR动态变化呈比较平坦的拱形,两端较低,中间较高;太阳辐射在第100~250 d即4—8月间相对较大,日平均PAR大于20 MJ·m-2·d-1,而这一段时间正是早晚稻生育期。气温的波动幅度比土壤温度大,气温和5 cm土壤温度明显呈现单峰曲线,在夏季(第150~250 d)日均温都超过了25 ℃,5 cm土温和气温具有极显著的线性相关,R2为0.861 9。从降雨量的柱状分布图可以看出2015年降雨较频繁,但雨量不大,年降雨量为1457 mm,降雨量幅度在0~50 mm之间,在第200~300 d内降雨次数较少,降雨量仅114 mm,而300~365 d降雨较多,达286 mm。

|
| 图 1 试验地气象与环境因子 Figure 1 Meteorological and environmental conditions in the study area |
如图 2所示,FC的月平均日变化清晰地显示了其季节性动态变化规律。在早晚稻生长季及冬闲期杂草生物量足够大时FC的月平均日变化总体呈“U”型单峰曲线(图 2b~图 2e)。1、2、11月和12月为冬季休闲且杂草群落较小,总体上表现为碳排放,FC的变化范围为-3.21~5 μmol·m-2·s-1,趋势相对平缓。3月和4月整体仍属于冬闲期间,但杂草群落足够大且温度升高,所以一方面白天的光合作用增强,另一方面白天和夜间的土壤呼吸和植物呼吸也增强。

|
| 图 2 碳通量的月平均日变化 Figure 2 Diurnal patterns of monthly averaged carbon flux |
5月至10月为早晚稻生育期,光辐射和气温以及水稻生物量都较高,白天的光合作用显著变强,呈现白天吸收CO2,晚上排放CO2的规律。日出半小时后CO2通量由正值(释放)转变为负值(吸收),随着太阳高度角的增大和太阳辐射的增强,FC不断增加,在中午12:00左右达到峰值,此后随着太阳辐射的降低,FC逐渐减小,傍晚19:00左右由负转变为正。5月份早稻刚移栽不久,处于生长前期,光合作用较弱,且土壤温度较高,土壤呼吸和水稻植株呼吸较强,白天FC集中在-2 μmol·m-2·s-1范围;6月为早稻主要生长季,光合能力强,白天FC集中在-8 μmol·m-2·s-1左右;7月早稻进入成熟期,叶片变黄,光合作用固碳能力降低,且土壤温度很高,土壤呼吸和水稻植株呼吸强,白天FC在0 μmol·m-2·s-1左右。8、9月为晚稻生长季,白天FC最高达-17.95 μmol·m-2·s-1,10月份晚稻进入成熟期,白天FC集中在-6 μmol·m-2·s-1左右。另外,5—10月水稻种植期间温度较高,尤其6—8月,土壤温度达到30 ℃,整个稻田生态系统夜间呼吸都很高,在10 μmol·m-2·s-1上下浮动。
2.2.2 NEE、GPP和Reco的季节变化如图 3所示,稻田NEE的季节变化呈明显的两个吸收期(NEE为负)和三个排放期(NEE为正)。一年两熟的双季稻模式使NEE在早稻和晚稻的生长季有两个明显的碳吸收期,早稻平均值为-0.58 g C·m-2,最大值出现在6月20日,为-1.77 g C·m-2·d-1,晚稻平均值为-1.28 g C·m-2·d-1最大值出现在9月19日,为-2.23 g C·m-2·d-1;在冬闲期的非作物生长期,存在两个碳排放期,平均值为2.68 g C·m-2·d-1。另外,在早晚稻的生长前期和后期,由于水稻的光合固碳能力较弱,也表现为碳排放,尤其是早稻收获后晚稻生长前期,呈现一个排放峰,NEE平均为7.14 g C·m-2·d-1。

|
| 图 3 NEE、GPP和Reco的季节性动态变化 Figure 3 Seasonal variation of daily and 7-day averaged NEE, GPP and Reco |
如图 3所示,冬闲田初期,稻田植物较少,GPP的量较低,随着气温升高以及杂草生长,GPP有少量增加。在早稻生长初期(4月22日—5月13日),由于水稻生物量及LAI较低,GPP也相对较低,在1~3 g C·m-2·d-1;随着LAI增大和温度的升高,GPP也逐渐上升,在7月1日达到最高值11.55 g C·m-2·d-1。此时土壤温度也高达30 ℃(图 1)。晚稻GPP与早稻有着同样的变化趋势,也是随着LAI增大而增加。
生态系统碳交换中的碳排放环节主要包括土壤呼吸和植物呼吸,这两者都受土壤温度的影响。如图 3中所示,Reco的变化幅度较大,为1.7~13.8 g C·m-2·d-1,其变化趋势与5 cm土壤温度的趋势一致(图 1),均为中间高、两端低。
2.2.3 “双季稻-冬闲田”生态系统碳收支图 4为碳交换的全年日累积动态变化,曲线的斜率表征碳交换每日的累积速率。NEE日累积曲线在双季稻生长期间表现为斜率变小。全年“双季稻-冬闲田”生态系统净碳交换的总和表现为向大气排放CO2,全年累积为778.4 g C·m-2,其中早稻时期和晚稻时期分别贡献147.7 g C·m-2和127.4 g C·m-2,冬闲时期则贡献了503.3 g C·m-2。与NEE相反,GPP的日累积动态变化曲线在双季稻生态系统表现为碳吸收时GPP每日的累积速率变大,全年稻田生态系统GPP为1 643.7 g C·m-2。Reco的每日累积速率受土壤温度影响,在1—3月以及11—12月期间累积速率较低,曲线较平缓;而在4—10月的水稻生长期间累积速率加大,曲线斜率较大,全年稻田生态系统Reco为2 425.8 g C·m-2。

|
| 图 4 NEE、GPP和Reco的日累积动态变化 Figure 4 Seasonal variation of cumulative NEE, GPP and Reco |
双季稻生态系统全年仅有88 d表现为碳吸收,且净吸收量不高。全年双季稻系统的Reco/GPP为1.48,因此,本研究中早晚稻通过光合作用固定的CO2低于碳排放总量,“双季稻-冬闲田”生态系统全年碳交换综合表现为碳源。
2.3 NEE的影响因子分析 2.3.1 5 cm土壤温度与夜间NEE的关系微生物与植物的新陈代谢通过呼吸作用向大气释放CO2,而一定范围内的高温会促进这种新陈代谢,进而增加碳排放。本研究分别选取冬闲期间、早稻生育期和晚稻生育期未进行插补的夜间CO2通量数据与5 cm土壤温度数据进行拟合,结果如图 5所示。各时期5 cm土壤温度均与夜间CO2通量拟合较好,有极显著的相关性(P < 0.000 1)。Q10值表征生态系统的土壤温度敏感性,值越高说明生态系统对土壤温度变化的响应越快。不同时期Q10值的大小为冬闲>晚稻>早稻,说明冬闲期间的稻田生态系统对温度的敏感性大于其他时期。

|
| 图 5 5 cm土壤温度与不同时期NEE的关系 Figure 5 Relationship between NEE in different stage and soil temperature at 5 cm |
白天NEE除了受气温和土壤温度影响,另一个主要影响因素是植物的光合作用强度。而植物光合作用强度与PAR有关。本研究分别选取早稻和晚稻种植期间未插补的晴朗天气数据,采用米氏方程进行拟合。如图 6所示,早稻和晚稻NEE与相应的PAR存在显著的相关性(P < 0.001),决定系数R2分别为0.53和0.39,即PAR分别解释了早稻53%的NEE变化和晚稻39%的NEE变化。早稻的Pmax与晚稻相当,但早稻白天的Rd低于晚稻,这与本文拟合时选取的NEE数据有关,早稻数据部分对应的土壤温度范围为18~24 ℃,晚稻部分对应土壤温度为23~30 ℃,如前所述,土壤温度与NEE呈指数相关,所以拟合方程中晚稻的Rd大于早稻。

|
| 图 6 PAR与NEE的关系 Figure 6 Relationship between PAR and NEE |
本研究表明,生态系统净碳交换(NEE)存在明显的日变化和季节性变化,具体体现为在作物生长季时生态系统从大气中吸收CO2,日变化明显,且总体呈“U”型曲线;而在非作物生长季时生态系统向大气排放CO2,日变化总体趋势平缓。这一特征在区域尺度上存在普遍性,差异只存在于NEE的变化幅度[24-27]。
白天农田生态系统碳交换在不同时间尺度上的变化主要受生态系统光合作用与呼吸作用强弱的影响,影响因子一般包括光合有效辐射、土壤温度、气体温度、植物生物量和叶面积指数等。光合有效辐射对植物光合作用强弱的影响制约着生态系统白天NEE的大小。本研究中,米氏方程能够较好地表示出在作物生长季中光合有效辐射和生态系统白天NEE的关系[28]。但是不同的生态系统类型,使用米氏方程进行模拟得到的相关关系大小存在较大差异,决定系数R2从0.3~0.9均有[7, 9, 28],本研究中决定系数也在这个范围内。差异的存在说明影响生态系统白天NEE大小除了光合有效辐射还有其他因子,比如气温、叶面积指数等,一方面是气温在影响光合作用的同时还影响植物以及土壤中微生物的呼吸;另一方面,叶面积指数大小影响光合作用的固碳量。而在夜间,不论是气温还是土壤温度,是影响夜间农田生态系统呼吸的一个主要环境因子,主要是通过影响土壤呼吸和植物呼吸发挥作用,而夜间无光合作用,生态系统呼吸大小即为NEE的大小。但对于选用气温还是土温能最好地解释夜间NEE的变化,目前没有定论,但通常的做法是采用5 cm土壤温度来进行解释[26, 28]。本研究中夜间NEE与土壤温度有极显著的正相关关系,Q10值代表生态系统的温度敏感性。在一年中,冬闲期间的Q10值比早晚稻期间的Q10值大,这与冬闲期间的时间跨度、温度幅度较大,且温度较低,土壤微生物对温度的响应更敏感有关[29]。另外,有研究表明,土壤水分含量对Q10值有显著影响,在土壤含水量低于40%时Q10值更大,而大于40%时则变小[30]。本研究中水稻种植期间土壤为淹水状态,含水量高,而冬闲期间不淹水,含水量低,所以冬闲期间的Q10值比早晚稻期间的Q10值大。
3.2 生态系统固碳量评估由于区域间气候与环境条件、种植制度、管理措施、土壤碳库大小和研究方法等存在较大差异,关于农田生态系统NEE的研究结果差异较大。有研究发现,稻田生态系统具有显著的碳汇效应,太湖流域典型稻麦轮作农田年NEE为-749.49~-785.38 g C·m-2 [24];淮河流域稻麦轮作系统年NEE为-626.92 g C·m-2 [25];华北平原的稻麦轮作系统年NEE为-197.6~-317.9 g C·m-2 [6];长三角地区的稻麦轮作系统年NEE达-769.6 g C·m-2 [26];双季稻农田生态系统的年NEE为-463.4 g C·m-2 [17]。而本研究中双季稻农田生态系统的NEE的累积变化表现为向大气中排放CO2,为778.4 g C·m-2,以一年的时间尺度看,本研究的“双季稻-冬闲田”生态系统在2015年整体表现为碳源。原因可能是涡度相关技术对夜间NEE的估算具有较大的不确定性,且通常都是低估的[31]。本研究中的夜间NEE冬闲期间在5 μmol·m-2·s-1左右,早晚稻种植期间在10 μmol·m-2·s-1左右,而其他使用涡度相关技术进行CO2通量监测的全年夜间NEE均值为1~4.5 μmol·m-2·s-1 [17, 24-26],所以本研究对生态系统夜间和白天的碳排放估算值相对较高(使用夜间NEE与土壤温度建立的van′t Hoff经验指数模型估算白天的碳排放)。但与他人用静态箱法估算的双季稻田碳排放量却相似。陈粲[32]在澳大利亚不同秸秆还田方式对稻田生态系统碳排放(Reco)的研究中,估算秸秆深埋方式下年排放达3500 g C·m-2,高于本研究的年碳排放量估算的2 425.8 g C·m-2。另外,刘兵[14]对鄂东南稻区的双季稻生态系统碳排放研究表明,不同种植方式下仅双季稻生长季稻田生态系统碳排放范围为4972~10 354 g C·m-2,更是远高于本研究估算的年碳排放。同样是稻麦轮作系统,黄明蔚[33]采用透明箱研究稻麦两季的NEE发现,无论在水稻生长季还是小麦生长季,均表现为碳排放,总量达928 g C·m-2。而且从全球碳平衡的角度看,每年由土壤释放的CO2等于或超过陆地生态系统的净初级生产力[34],这意味着土壤排放的CO2量超过了植物通过光合作用固定的CO2。因此,提高农田土地在时间和空间上的植被覆盖率,将有可能使植物光合作用固定的碳超过土壤呼吸释放的碳。
前期本项目组采用透明静态箱法研究冬闲田种植黑麦草和紫云英的白天的净碳交换,结果表明冬季种植黑麦草和紫云英能提高稻田生态系统白天的固碳量,至早稻种植前,种植紫云英处理白天固碳总计381 g C·m-2,种植黑麦草处理为300 g C·m-2 [35]。表明持续时间长达5个半月的稻田冬闲期具有较高的固碳潜力,充分利用将很可能有效提高稻田全年生态系统固碳能力。当前国内外利用冬闲田种植作物的生态系统全年净碳交换研究还鲜见报道,因此,研究全年覆盖作物的农田生态系统净碳交换对于理解和评价冬闲田或其他类型农闲田的充分利用在农业生产和固碳减排效应上的作用具有重要意义。
4 结论(1)双季稻生态系统碳交换存在明显的日变化和季节变化,而导致变化的直接原因是生态系统处于生长季还是非生长季。在生长季中,水稻种植期间白天的净碳交换受光合有效辐射的影响显著;全年夜间的净碳交换受5 cm土壤温度的显著影响。土温低时的冬闲时期土壤温度敏感性高于土温高时的双季稻生长季。
(2)该生态系统全年净碳交换为778.4 g C·m-2,总初级生产力为1 643.7 g C·m-2,碳排放为2 425.8 g C·m-2,其中冬闲田期间占64.7%。可见水稻光合作用固定了大量CO2,为稻田生态系统固碳减排做出了巨大贡献。但由于双季稻这一种植模式中有长达4~6个月的闲置期,而冬闲期间具有较高的碳排放,导致该种植模式下水稻生态系统全年整体表现为碳排放。因此,如能充分利用好冬闲田以及提高双季稻光合作用效率,“双季稻-冬闲田”生态系统有可观的固碳减排潜力。
| [1] |
IPCC. Climate change 2013: The physical science basis. Contribution of working group Ⅰ to the fifth assessment report of the Intergovernmental Panel on Climate Change[M]. Cambridge and New York: Cambridge University Press, 2013: 1535.
|
| [2] |
李国栋, 张俊华, 陈聪, 等. 气候变化背景下中国陆地生态系统碳储量及碳通量研究进展[J]. 生态环境学报, 2013, 22(5): 873-878. LI Guo-dong, ZHANG Jun-hua, CHEN Cong, et al. Research progress on carbon storage and flux in different terrestrial ecosystem in China under global climate change[J]. Ecology and Environmental Sciences, 2013, 22(5): 873-878. |
| [3] |
Soegaard H, Jensen N O, Boegh E, et al. Carbon dioxide exchange over agricultural landscape using eddy correlation and footprint modeling[J]. Agricultural and Forest Meteorology, 2003, 114(3/4): 153-173. |
| [4] |
梁涛, 李荣平, 吴航, 等. 玉米农田生态系统CO2通量的动态变化[J]. 气象与环境学报, 2012, 28(3): 49-53. LIANG Tao, LI Rong-ping, WU Hang, et al. Dynamics of carbon dioxide flux in a maize agroecosystem[J]. Journal of Meteorology and Environment, 2012, 28(3): 49-53. |
| [5] |
薛红喜, 李峰, 李琪, 等. 基于涡度相关法的中国农田生态系统碳通量研究进展[J]. 南京信息工程大学学报:自然科学版, 2012, 4(3): 226-232. XUE Hong-xi, LI Feng, LI Qi, et al. Research progress on carbon flux over agro-ecosystem based on the eddy covariance method in China[J]. Journal of Nanjing University of Information Science & Technology(Natural Science Edition), 2012, 4(3): 226-232. |
| [6] |
李俊, 于强, 孙晓敏, 等. 华北平原农田生态系统碳交换及其环境调控机制[J]. 中国科学D辑:地球科学, 2006, 36: 210-223. LI Jun, YU Qiang, SUN Xiao-min, et al. Carbon dioxide exchange and the mechanism of environmental control in a farmland ecosystem in North China Plain[J]. Science in China Series D:Earth Sciences, 2006, 36(Suppl 1): 210-223. |
| [7] |
于贵瑞, 张雷明, 孙晓敏. 中国陆地生态系统通量观测研究网络(ChinaFLUX)的主要进展及发展展望[J]. 地理科学进展, 2014, 33(7): 903-917. YU Gui-rui, ZHANG Lei-ming, SUN Xiao-min. Progresses and prospects of Chinese terrestrial ecosystem flux observation and research net work(ChinaFLUX)[J]. Progress in Geography, 2014, 33(7): 903-917. DOI:10.11820/dlkxjz.2014.07.005 |
| [8] |
林同保, 王志强, 宋雪雷, 等. 冬小麦农田二氧化碳通量及其影响因素分析[J]. 中国生态农业学报, 2008, 16(6): 1458-1463. LIN Tong-bao, WANG Zhi-qiang, SONG Xue-lei, et al. CO2 flux and impact factors in winter wheat field ecosystem[J]. Chinese Journal of Eco-Agriculture, 2008, 16(6): 1458-1463. |
| [9] |
许琰, 周石硚, 晋绿生, 等. 新疆北部覆膜滴灌棉田的碳交换日、生长季变化特征[J]. 干旱区地理, 2013, 36(3): 441-449. XU Yan, ZHOU Shi-qiao, JIN Lü-sheng, et al. Diurnal and seasonal variation of carbon dioxide exchange over a film-mulched cotton field under drip irrigation in Northern Xinjiang[J]. Arid Land Geography, 2013, 36(3): 441-449. |
| [10] |
苏荣瑞, 刘凯文, 耿一风, 等. 江汉平原稻田冠层CO2通量变化特征及其影响因素分析[J]. 长江流域资源与环境, 2013, 22(9): 1214-1220. SU Rong-rui, LIU Kai-wen, GENG Yi-feng, et al. Characteristics of canopy CO2 flux over paddy ecosystem in Jianghan Plain and its influence factors analysis[J]. Resources and Environment in the Yangtze Basin, 2013, 22(9): 1214-1220. |
| [11] |
韩广轩, 周广胜, 许振柱. 玉米农田生态系统土壤呼吸作用季节动态与碳收支初步估算[J]. 中国生态农业学报, 2009, 17(5): 874-879. HAN Guang-xuan, ZHOU Guang-sheng, XU Zhen-zhu. Seasonal dynamics of soil respiration and carbon budget of maize(Zea mays L.) farmland ecosystem[J]. Chinese Journal of Eco-Agriculture, 2009, 17(5): 874-879. |
| [12] |
梁尧, 韩晓增, 乔云发, 等. 小麦-玉米-大豆轮作下黑土农田土壤呼吸与碳平衡[J]. 中国生态农业学报, 2012, 20(4): 395-401. LIANG Yao, HAN Xiao-zeng, QIAO Yun-fa, et al. Soil respiration and carbon budget in black soils of wheat-maize-soybean rotation system[J]. Chinese Journal of Eco-Agriculture, 2012, 20(4): 395-401. |
| [13] |
国家统计局. 中国统计年鉴[EB/OL]. [2017-08-29]. http://data.stats.gov.cn/easyquery.htm?cn=C01. National Bureau of Statistics. Statistical yearbook of china[EB/OL]. [2017-08-29]. http://data.stats.gov.cn/easyquery.htm?cn=C01. |
| [14] |
刘兵. 双季稻不同耕作与种植方式对产量和农田碳排放的影响[D]. 武汉: 华中农业大学, 2013. LIU Bing. Effect of cultivation methods and planting patterns on double cropping yield and carbon fluxes emission from the paddy field[D]. Wuhan: Huazhong Agricultural University, 2013. |
| [15] |
秦晓波, 李玉娥, WANGHong, 等. 生物质炭添加对华南双季稻田碳排放强度的影响[J]. 农业工程学报, 2015, 31(5): 226-234. QIN Xiao-bo, LI Yu-e, WANG Hong, et al. Impact of biochar amendment on carbon emissions intensity in double rice field in South China[J]. Transactions of the Chinese Society of Agricultural Engineering, 2015, 31(5): 226-234. |
| [16] |
马艳芹, 钱晨晨, 孙丹平, 等. 施氮水平对稻田温室气体排放的影响[J]. 农业工程学报, 2016, 32(增刊2): 128-134. MA Yan-qin, QIAN Chen-chen, SUN Dan-ping, et al. Effect of nitrogen fertilizer application on greenhouse gas emissions from soil in paddy field[J]. Transactions of the Chinese Society of Agricultural Engineering, 2016, 32(Suppl 2): 128-134. |
| [17] |
王尚明, 胡继超, 吴高学, 等. 亚热带稻田生态系统CO2通量特征分析[J]. 环境科学学报, 2011, 31(1): 217-224. WANG Shang-ming, HU Ji-chao, WU Gao-xue, et al. Characteristics analysis of CO2 flux from a rice paddy ecosystem in a subtropical region[J]. Acta Scientiae Circumstantiae, 2011, 31(1): 217-224. |
| [18] |
宋涛. 三江平原生态系统CO2通量的长期观测研究[D]. 南京: 南京信息工程大学, 2007. SONG Tao. Long-term observation of CO2 fluxes in the ecosystem of Sanjiang Plain[D]. Nanjing: Nanjing University of Information Science & Technology, 2007. |
| [19] |
Xu L K, Baldocchi D B. Seasonal variation in carbon dioxide exchange over a Mediterranean annual grassland in California[J]. Agricultural and Forest Meteorology, 2004, 123: 79-96. DOI:10.1016/j.agrformet.2003.10.004 |
| [20] |
Katul G, Lai C T, Schäfer K, et al. Multiscale analysis of vegetation surface fluxes:From seconds to years[J]. Advances in Water Resources, 2001, 24(9/10): 1119-1132. |
| [21] |
张弥, 温学发, 于贵瑞, 等. 二氧化碳储存通量对森林生态系统碳收支的影响[J]. 应用生态学报, 2010, 21(5): 1201-1209. ZHANG Mi, WEN Xue-fa, YU Gui-rui, et al. Effects of CO2 storage fluxon carbon budget of forest ecosystem[J]. Chinese Journal of Applied Ecology, 2010, 21(5): 1201-1209. |
| [22] |
陈云飞, 江洪, 周国模, 等. 人工高效经营雷竹林CO2通量估算及季节变化特征[J]. 生态学报, 2013, 33(11): 3434-3444. CHEN Yun-fei, JIANG Hong, ZHOU Guo-mo, et al. Estimation of CO2 fluxes and its seasonal variations from the effective management Lei bamboo(Phyllostachys violascens)[J]. Acta Ecologica Sinica, 2013, 33(11): 3434-3444. |
| [23] |
Schmidt M, Reichenau T G, Fiener P, et al. The carbon budget of a winter wheat field:An eddy covariance analysis of seasonal and inter-annual variability[J]. Agricultural and Forest Meteorology, 2012, 165: 114-126. DOI:10.1016/j.agrformet.2012.05.012 |
| [24] |
徐昔保, 杨桂山, 孙小祥. 太湖流域典型稻麦轮作农田生态系统碳交换及影响因素[J]. 生态学报, 2015, 35(20): 6655-6665. XU Xi-bao, YANG Gui-shan, SUN Xiao-xiang. Analysis of net ecosystem CO2 exchange(NEE) in the rice-wheat rotation agroecosystem of the Lake Taihu Basin, China[J]. Acta Ecologica Sinica, 2015, 35(20): 6655-6665. |
| [25] |
李琪, 胡正华, 薛红喜, 等. 淮河流域典型农田生态系统碳通量变化特征[J]. 农业环境科学学报, 2009, 28(12): 2545-2550. LI Qi, HU Zheng-hua, XUE Hong-xi, et al. Variation of net ecosystem carbon flux over typical agro-ecosystem in Huaihe River Basin[J]. Journal of Agro-Environment Science, 2009, 28(12): 2545-2550. DOI:10.3321/j.issn:1672-2043.2009.12.017 |
| [26] |
孙小祥, 常志州, 杨桂山, 等. 长三角地区稻麦轮作生态系统净碳交换及其环境影响因子[J]. 中国生态农业学报, 2015, 23(7): 803-811. SUN Xiao-xiang, CHANG Zhi-zhou, YANG Gui-shan, et al. Characteristics of net ecosystem exchange and environmental factors of rice-wheat rotation system in the Yangtze River Delta of China[J]. Chinese Journal of Eco-Agriculture, 2015, 23(7): 803-811. |
| [27] |
张法伟, 李英年, 曹广民, 等. 青海湖北岸高寒草甸草原生态系统CO2通量特征及其驱动因子[J]. 植物生态学报, 2012, 36(3): 187-198. ZHANG Fa-wei, LI Ying-nian, CAO Guang-min, et al. CO2 fluxes and their driving factors over alpine meadow grassland ecosystems in the northern shore of Qinghai Lake, China[J]. Chinese Journal of Plant Ecology, 2012, 36(3): 187-198. |
| [28] |
朱咏莉, 吴金水, 童成立, 等. 稻田CO2通量对光强和温度变化的响应特征[J]. 环境科学, 2008, 29(4): 1040-1044. ZHU Yong-li, WU Jin-shui, TONG Cheng-li, et al. Responses of CO2 fluxes to light intensity and temperature in rice paddy field[J]. Environmental Science, 2008, 29(4): 1040-1044. |
| [29] |
盛浩, 杨玉盛, 陈光水, 等. 土壤异养呼吸温度敏感性(Q10)的影响因子[J]. 亚热带资源与环境学报, 2006, 1(1): 74-82. SHENG Hao, YANG Yu-sheng, CHEN Guang-shui, et al. Controls on the temperature sensitivity of soil heterotrophic respiration:Q10 variability and analysis[J]. Journal of Subtropical Resources and Environment, 2006, 1(1): 74-82. |
| [30] |
Han G X, Xing Q H, Yu J B, et al. Agricultural reclamation effects on ecosystem CO2 exchange of a coastal wetland in the Yellow River Delta[J]. Agriculture, Ecosystems and Environment, 2014, 196(1793): 187-198. |
| [31] |
朱治林, 孙晓敏, 温学发, 等. 中国通量网(ChinaFLUX)夜间CO2涡度相关通量数据处理方法研究[J]. 中国科学D辑:地球科学, 2006, 36(增刊1): 34-44. ZHU Zhi-lin, SUN Xiao-min, WEN Xue-fa, et al. The study of eddy covariance flux data processing of China FLUX network[J]. Science in China Series D:Earth Sciences, 2006, 36(Suppl 1): 34-44. |
| [32] |
陈粲. 不同秸秆还田处理下农田温室气体排放的DNDC模型模拟及模型适应性研究[D]. 南京: 南京农业大学, 2014. CHEN Can. The DNDC model simulation of agriculture green house gases emission and the model adaptability research under different straw retention methods[D]. Nanjing: Nanjing Agricultural University, 2014. |
| [33] |
黄明蔚. 稻麦轮作模式农田生态系统温室气体排放及机制研究[D]. 上海: 华东师范大学, 2007. HUANG Ming-wei. Emission progress of green house gases and its mechanism from the paddy-wheat rotation agro-ecosystem[D]. Shanghai: East China Normal University, 2007. |
| [34] |
方精云, 朴世龙, 赵淑清. CO2失汇与北半球中高纬度陆地生态系统的碳汇[J]. 植物生态学报, 2001, 25(5): 594-602. FANG Jing-yun, PIAO Shi-long, ZHAO Shu-qing. The carbon sink:The role of the middle and high latitudes terrestrial ecosystems in the Northern Hemisphere[J]. Acta Phytoecologica Sinica, 2001, 25(5): 594-602. |
| [35] |
魏甲彬, 周玲红, 徐华勤, 等. 南方种养结合模式对冬季稻田净碳交换和不同土层活性碳氮转化的影响[J]. 草业学报, 2017, 26(7): 138-146. WEI Jia-bin, ZHOU Ling-hong, XU Hua-qin, et al. Effects of forage planting and chickens on net carbon exchange and transformation of soil active carbon and nitrogen at different layers in paddy fields in South China in winter[J]. Acta Prataculturae Sinica, 2017, 26(7): 138-146. DOI:10.11686/cyxb2017020 |