 2019, Vol. 38
2019, Vol. 38



2. 安徽省养分循环与资源环境省级实验室/安徽省农业科学院土壤肥料研究所, 合肥 230031;
3. 中国科学院大学, 北京 100049
2. Laboratory of Nutrient Cycling, Resources and Environment of Anhui, Institute of Soil and Fertilizer, Anhui Academy of Agricultural Sciences, Hefei 230031, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
水稻是中国乃至亚洲的主要种植作物之一, 约占世界粮食种植面积的1/3, 养活了世界一半以上的人口, 对保障世界粮食安全举足轻重[1-2]。人类活动导致的大气CO2浓度([CO2])和温度的不断升高对水稻的生长发育产生了重要影响[3]。
已有大量研究表明, 高[CO2]提高了水稻的光合速率, 促进碳水化合物的合成, 增加有效分蘖数, 提高了水稻产量[4-7]。但长期高[CO2]下水稻存在光合下调现象, 限制了产量的增幅[7]。水稻对温度比较敏感, 其中开花期和灌浆期受温度影响最大, 当开花期气温高于34 ℃, 可阻止花粉膨大, 迫使孢子膜破裂[8], 导致不育籽粒增加[9], 又因呼吸作用增加了碳水化合物的消耗导致水稻减产[5-6, 10]。产量构成是揭示[CO2]和温度对水稻的影响较直接的因素。万运帆等[11]研究表明, 增温或[CO2]和温度同时增加条件下早稻增产主要与穗数和穗粒数的增加有关, 而空瘪率的增加限制了产量的增幅。Wang等[6]认为高[CO2]和高温下枝梗数及其对应着生的籽粒数减少导致单位面积总颖花数降低, 是水稻减产的主要原因。水稻颖花开花的先后顺序为上部、中部、下部[12], 高温导致水稻灌浆期籽粒上部、中部、下部的结实率逐渐降低[13]。位于水稻籽粒上部的强势粒充实度好、粒质量大、结实率高, 而位于下部的弱势粒充实度差、粒质量小、结实率低[14]。在未来[CO2]和温度相伴升高的条件下, 水稻不同粒位籽粒粒质量将会如何变化, 对籽粒充实度的影响如何, 对水稻产量的影响又如何, 尚不明确, 亟待研究。
相对于封闭或半封闭的气室模拟未来[CO2]和温度相伴升高的环境[11, 15], 本研究利用先进的开放自然空间CO2升高平台(FACE) [16], 采用热水管道为水稻冠层增温的方式建成的增温-FACE平台(T-FACE), 能更真实地模拟2050年大气环境[5]。因此, 本研究基于T-FACE平台, 以优质水稻南粳9108为材料, 研究[CO2]和温度升高对水稻不同粒位粒质量的影响, 旨在明确未来[CO2]和温度相伴升高下水稻籽粒充实度变化规律, 为预测水稻产量响应未来气候变化提供理论依据。
1 材料与方法 1.1 试验地区及平台概况试验于2015年和2016年在中国稻田T-FACE研究技术平台上进行。平台位于江苏省扬州市江都区小纪镇良种场试验田(119°42′ 00″ E, 32°35′ 05″ N)。试验田所在地区年均降水量980 mm左右, 年均蒸发量大于1100 mm, 年平均温度约14.9 ℃, 年日照时间大于2100 h, 年平均无霜期220 d, 耕作方式为水稻-冬小麦轮作。
试验田土壤类型为砂壤土, 土壤理化性质为:有机质18.4 g·kg-1, 全氮1.45 g·kg-1, 全磷0.63 g·kg-1, 全钾14.0 g·kg-1, 速效磷10.1 mg·kg-1, 速效钾70.5 mg· kg-1, 容重1.16 g·cm-3, pH 7.2。
平台共有3个FACE试验圈和3个对照(Ambient)圈, 各圈间距大于90 m。FACE圈是直径12 m正八边形, 平台运行时通过FACE圈周围的管道向中心喷射纯CO2气体, 并在FACE和Ambient圈中特定位置放置长、宽分别为2.70、0.75 m(7个)的热水增温管道, 以热辐射形式向增温区域进行增温处理。
利用计算机网络对平台[CO2]和水稻冠层温度进行监测和控制, 根据[CO2]、风向、风速、作物冠层高度的[CO2]和温度自动调节CO2气体的释放速度和方向以及增温管道中热水流速, 使水稻主要生育期FACE圈内[CO2]保持比大气环境高200 μmol·mol-1, 所有圈内增温区域水稻冠层空气温度比大气环境温度高1 ℃左右。对照田块没有安装FACE管道, 所有田块非增温区域没有安装热水增温管道, 其余环境条件与自然状态一致[5, 16]。为监测增温装置为大气实际增温的情况, 在FACE和Ambient圈增温与非增温区域, 距水稻冠层上方20 cm处安装SI-111红外温度传感仪(Campbell公司, 美国)实时记录气温, 每1 min记录一次。
1.2 试验材料与栽培试验于2015年和2016年水稻季进行, 供试品种为南粳9108。苗龄为4周的秧苗, 两年均于6月17日移栽, 种植密度为24穴·m-2, 每穴2苗, 秧苗均为1蘖苗。采用复合肥(有效成分N:P2O5:K2O=15%:15%: 15%)和尿素(含N 46.2%)配合施用。施N总量为22.5 g·m-2, 基肥、分蘖肥和穗肥的施用比例为40%、30%和30%。施P、K量均为9 g·m-2, 作基肥施用。2015年10月26日和2016年11月2日收获。其他田间管理如水分、病虫害等同大田一致。
1.3 试验设计试验设置4个处理, 设大气背景[CO2](Ambient [CO2], 约390 μmol·mol-1)和高[CO2] (Elevated [CO2], 约590 μmol·mol, 比大气背景[CO2]高200 μmol·mol-1) 2个水平。裂区为温度处理, 设大气环境温度(Ambient temperature)和高温(Elevated temperature, 比环境温度高1 ℃) 2个水平。具体为环境[CO2]和温度处理(Ambient)、环境[CO2]和温度升高处理(eT)、[CO2]升高和环境温度处理(eC)、[CO2]和温度均升高处理(eT+eC)。
2015年平台CO2熏气时间为6月25日至10月19日, 增温时间为7月1日至10月19日;2016年CO2熏气时间为6月25日至10月19日, 增温时间为6月28日至11月2日。FACE圈每日熏气和增温处理时间为日出至日落。
1.4 样品采集与分析水稻齐穗期后, 每个处理选具有代表性的植株8穴, 将长势一致的所有单穗进行挂牌, 在灌浆初期、灌浆中期、灌浆后期、成熟期按照不同粒位收获籽粒。
按照生育期每个处理采2穴样品, 数其挂牌穗数, 按平均法将稻穗分为上、中、下3个部位, 将位于穗上部第一枝梗上的第1、2、3粒合并, 为强势粒, 穗下部一次枝梗上着生于二次枝梗上的籽粒(除顶部第一粒除外)为弱势粒[17], 其余籽粒为中势粒。2穴中挂牌穗质量的强、中、弱势粒分别合并为单独样本, 在数完穗粒数和强、中、弱势粒粒数后, 80 ℃烘箱烘干至恒质量, 分别称质量, 根据其挂牌穗数计算单穗籽粒质量, 强、中、弱势粒穗粒质量及其对应千粒质量。计算公式为:
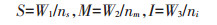
|
式中:S表示强势粒千粒质量, g;M表示中势粒千粒质量, g;I表示弱势粒千粒质量, g;W1、W2、W3分别表示2穴水稻强、中、弱势粒粒质量, g;ns、nm、ni分别表示2穴水稻强、中、弱势粒粒数。
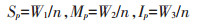
|
式中:Sp表示强势粒穗粒质量, g;Mp表示中势粒穗粒质量, g;Ip表示弱势粒穗粒质量, g;n表示2穴水稻有效穗数。
灌浆期采样时间:2015年的灌浆初期、灌浆中期、灌浆后期分别于9月11日、9月24日、10月3日采样, 2016年对应的分别于9月11日、9月22日、10月5日采样。
收获期, 每个处理数50穴调查有效穗数, 实收测产, 取5穴样品进行常规考种。先统计5穴样品的有效穗数, 再通过人工脱粒。将刘建丰等[18]的水稻籽粒充实状况指标测定方法改进后分离饱瘪粒(饱粒、半饱粒、空瘪粒)。具体操作方法为先后通过蒸馏水和80%酒精进行漂洗:准备好一装水的容器, 将待测水稻籽粒倒入水中, 搅拌4 min再静止1 min, 沉下去的为饱粒;将其余未沉下去的水稻籽粒全部用网筛捞出, 烘干后再放入80%酒精中, 搅拌4 min再静止1 min, 沉入80%酒精的籽粒为半饱粒, 未沉下去的籽粒为空瘪粒。将分离后饱粒、半饱粒、空瘪粒在80 ℃下烘干至恒质量, 分别称总质量, 每个饱粒和空瘪粒样品分别称取3个30.00 g和2.10 g的样本, 分别数样本的饱粒和空瘪粒的粒数, 半瘪粒全部数粒数, 计算总粒数、饱粒千粒质量和结实率。计算公式为:
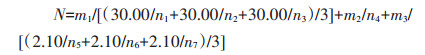
|
式中:N表示总粒数;m1、m2、m3分别表示总的饱粒、半饱粒、空瘪粒烘干质量, g;n1、n2、n3分别表示3个30.00 g样本的饱粒粒数;n4表示半饱粒粒数;n5、n6、n7分别表示3个2.10 g样本的空瘪粒粒数。饱粒千粒质量计算式为:
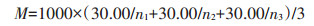
|
式中:M表示饱粒千粒质量。
结实率计算公式为:
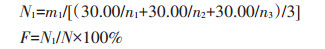
|
式中:F表示结实率, %;N1表示饱粒数;N表示总粒数。
1.5 数据处理与分析应用Excel 2010进行数据处理和图表绘制及统计分析。采用SPSS 20以一般线性模型进行方差分析, 以Tukey HSD法进行多重比较, 显著水平设P < 0.01、P < 0.05、P < 0.1和P>0.1, 分别用**、*、+和ns表示。图表中数据为平均值±标准误差。
2 结果与分析 2.1 2015年和2016年水稻季田间气象数据从图 1可知, 2016年水稻季6—8月平均气温比2015年同时期气温高1.5 ℃以上, 其中7月份高5 ℃。2016年稻季高温超过34 ℃的天数比2015年多10 d。2015年和2016年水稻季降雨天数分别为46、56 d, 降雨总量分别为972.4、1 109.5 mm, 其中2016年9月份至收获共降雨28 d, 降雨量达564.3 mm, 降雨天数比2015年多21 d, 降雨量多388.1 mm (图 2)。

|
图 1 水稻生长季的大气温度 Figure 1 Daily temperature in rice growing season |

|
图 2 水稻生长季的降雨量 Figure 2 Daily precipitation in rice growing season |
从图 3可以看出, 利用红外温度传感仪测定, 2015年和2016年稻季环境[CO2]和温度升高处理(eT)气温比环境[CO2]和温度处理(Ambient)分别平均增加了1.11 ℃和1.04 ℃, [CO2]和温度均升高处理(eT+eC)气温比Ambient分别平均增加了1.38 ℃和1.39 ℃, 达到试验设计的增温设备使水稻冠层气温增高1 ℃左右的目标。大气[CO2]升高200 μmol·mol-1导致气温增加, 数据显示, 2015年和2016年稻季[CO2]升高和环境温度处理(eC)气温比Ambient分别平均增加了0.47 ℃和0.45 ℃。

|
图 3 水稻生长季红外温度传感仪检测的不同处理的气温差异 Figure 3 Temperature was monitored by an infrared sensor for the treatments in 2015 and 2016 in rice growing season |
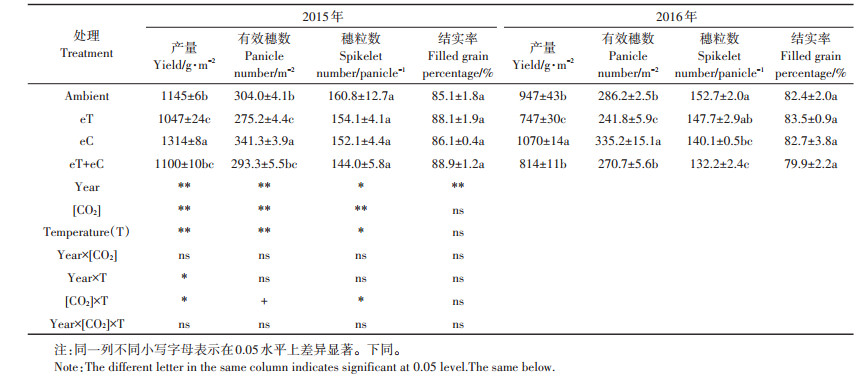
由表 1可知, 两年结果差异明显, 但不同处理间差异相似。高[CO2]增加了水稻产量、有效穗数和减少了穗粒数, 高温减少了水稻产量、有效穗数和穗粒数, [CO2]和温度对产量(P < 0.05)、有效穗数(P < 0.1)和穗粒数(P < 0.05)有不同程度的交互作用, 但对水稻结实率影响不大。与Ambient相比, 2015年eC产量增加了14.8%, 有效穗数增加了12.2%, 2016年eC产量和有效穗数分别增加了12.9%和17.1%;2015年和2016年eT产量分别减少了8.6%和21.1%;eT+eC产量分别减少了4.0%和14.0%, 相应的有效穗数分别减少3.5%和5.4%。两年中eT+eC比eC平均减产了20.1%, eT比Ambient平均减产14.8%, 前者减幅明显高于后者, 表明[CO2]升高下, 增温对水稻产量的负效应加剧。
|
|
表 1 大气CO2浓度和温度升高下水稻产量及其构成 Table 1 Yield and yield components of rice under elevated [CO2] and warmed temperature |
从表 2可知, 两年水稻千粒质量(饱粒、强、中、弱势粒千粒质量)差异明显。收获期, 2015年水稻千粒质量显著高于2016年(P < 0.01), 饱粒、强、中势粒千粒质量分别增加了7.6%~11.3%、10.2%~19.1%、10.0%~15.1%, 弱势粒千粒质量除了Ambient减少了6.0%, eT、eC、eT + eC分别增加了19.2%、24.8%、23.3%。并且, 2015年强势粒千粒质量比饱粒质量的增幅高于2016年。
|
|
表 2 大气CO2浓度和温度升高下水稻千粒质量(g) Table 2 1000 grain weight of rice under elevated [CO2] and warmed temperature (g) |

[CO2]升高、温度增加均未影响水稻饱粒千粒质量和强势粒千粒质量, 但[CO2]升高增加了中、弱势粒千粒质量, 温度升高降低了中势粒千粒质量(表 2)。与Ambient和eC比, 2015年eT+eC中势粒千粒质量降低略有下降, 2016年降幅较明显, 分别为4.8%和5.7%。
2.4 大气CO2浓度和温度升高下强、中、弱势粒穗粒质量在不同生育期和处理下, 2015年水稻单穗粒质量高于2016年(图 4)。收获期, 2015年水稻中势粒穗粒质量仍高于2016年, 但强、弱势粒穗粒质量低于2016年(图 5、图 6、图 7)。具体表现为, 收获期2015年水稻单穗粒质量和中势粒穗粒质量比2016年增加了20.2%~24.5%和32.1%~37.5%, 强、弱势粒穗粒质量比2016年减少了10.8%~16.7%和5.2%~25.9%。

|
EGFS:灌浆初期;MGFS:灌浆中期;LGFS:灌浆后期;Harvest:收获期。不同小写字母表示在0.05水平上差异显著。下同 EGFS, MGFS, LGFS are the abbreviations of"early grain filling stage", "mid grain filling stage", "late grain filling stage".The different letters in the figure indicate significant at 0.05 level. The same below 图 4 大气CO2浓度和温度升高下水稻单穗质量 Figure 4 Grain weight per panicle of rice under elevated [CO2] and warmed temperature |

|
图 5 大气CO2浓度和温度升高下水稻强势粒穗粒质量 Figure 5 Superior grain weight per panicle of rice under elevated [CO2] and warmed temperature |

|
图 6 大气CO2浓度和温度升高下水稻弱势粒穗粒质量 Figure 6 Inferior grain weight per panicle of rice under elevated [CO2] and warmed temperature |

|
图 7 大气CO2浓度和温度升高下水稻中势粒穗粒质量 Figure 7 Medium strength grain weight per panicle of rice under elevated [CO2] and warmed temperature |
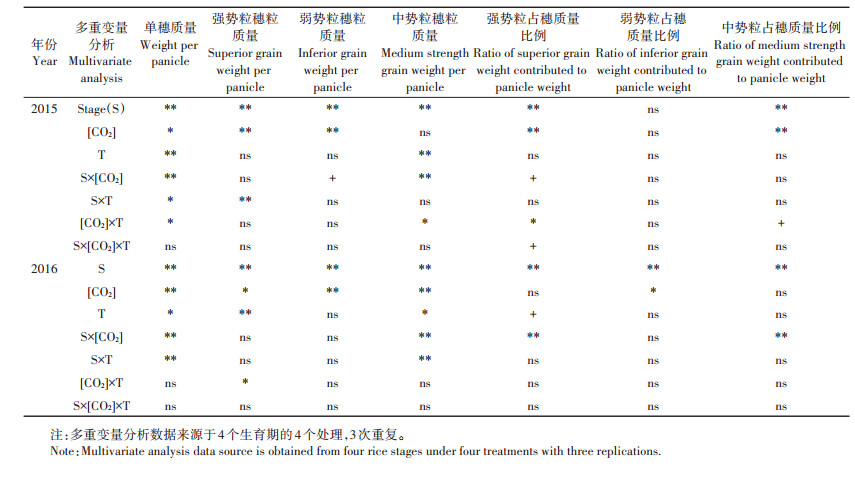
随着水稻生长发育, 单穗粒质量增幅明显, [CO2]、温度及二者交互作用对单穗粒质量的影响与水稻生育期和年份有关(表 3、图 4)。由于[CO2]和温度升高促进水稻生长发育和缩短生育期, 导致与Ambient相比, 在灌浆初期eT+eC两年的单穗粒质量增加明显, 而在灌浆中期和后期, eT+eC两年的单穗粒质量差异不大。收获期, 单独升高[CO2]和温度或二者同时升高均降低了单穗粒质量(表 4、图 2), 与Ambient相比, 2015年和2016年单穗粒质量eC分别减少了3.1%和4.2%, eT分别减少了4.3%和6.5%, eT+eC分别减少了5.5%和8.8%。
|
|
表 3 单穗质量、强势粒穗粒质量、弱势粒穗粒质量、中势粒穗粒质量及其占穗质量比例的多变量分析 Table 3 Grain weight, superior grain weight, inferior grain weight, medium strength grain weight per panicle, and its ratio contributed to panicle weight, respectively |
|
|
表 4 收获期单穗质量、强势粒穗粒质量、弱势粒穗粒质量、中势粒穗粒质量及其占穗质量比例的多变量分析 Table 4 Grain weight, superior grain weight, inferior grain weight, medium strength grain weight per panicle, and its ratio contributed to panicle weight in harvest, respectively |
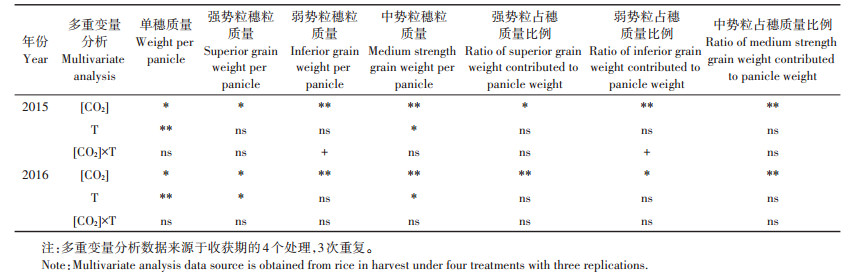
[CO2]升高增加了强、弱势粒穗粒质量, 降低了中势粒穗粒质量(表 3、表 4, 图 5~图 7)。收获期, 与Ambient相比, 2015年和2016年eC强势粒穗粒分别增加了14.2%和13.6%, 弱势粒穗粒质量分别增加了41.9%和10.8%, 中势粒穗粒质量分别减少了7.9%和9.2%。
增温对籽粒穗粒质量的影响与水稻生育期和籽粒粒位有关(表 3、表 4, 图 4~图 7)。灌浆初期, 增温不同程度促进了水稻强、中、弱势粒穗粒质量增加。收获期, 增温减少了强、中势粒穗粒质量, 对弱势粒穗粒质量的影响不明显。与Ambient相比, 2015年和2016年eT中势粒穗粒质量分别减少了5.1%和6.9%, 高于强势粒的降幅。
[CO2]和温度升高使强、弱势粒穗粒质量增加, 中势粒穗粒质量减少(表 3、表 4, 图 5~图 7), 与高[CO2]对强、中、弱势粒穗粒质量影响的结果相似, 增温总体上降低了高[CO2]对强、弱势粒粒质量的正效应, 加剧了其对中势粒穗粒质量的负效应。收获期, 与Ambient相比, 2015年和2016年eT+eC强势粒穗粒质量分别增加了8.0%和0.8%, 弱势粒穗粒质量分别增加了33.6%和13.8%, 中势粒穗粒质量分别减少了9.5%和11.3%。
2.5 大气CO2浓度和温度升高下强、中、弱势粒占穗质量比例变化在水稻灌浆成熟期过程中, 高[CO2]增加了水稻强势粒占穗质量比例, 增温增加了弱势粒占穗质量比例, 分别在2015年(P < 0.01)和2016年(P < 0.05)达到了显著水平, 同时, 在2015年高[CO2]显著降低了中势粒占穗质量比例, [CO2]和温度升高对强、中、弱势粒占穗质量比例无交互影响(表 3)。进一步分析, 收获期, 2015年和2016年高[CO2]对水稻强、中、弱势粒占穗质量比例的影响均达显著水平, 而增温及[CO2]和温度同时升高对其影响不明显(表 4)。与Ambient相比, 2015年和2016年收获期eC强势粒占穗质量比例分别增加了17.9%和18.7%, 而eT+eC强势粒占穗质量比例增幅下降, 分别增加了14.4%和10.5%;eC弱势粒占穗质量比例分别增加了46.4%和15.7%, 而eT+eC弱势粒占穗质量比例分别增加了41.4%和24.8%;eC中势粒占穗质量比例分别下降5.0%和5.2%, 而eT+eC中势粒占穗质量比例分别下降4.2%和4.7% (图 8)。

|
图 8 大气CO2浓度和温度升高下水稻强、中、弱势粒质量占穗质量比例 Figure 8 Ratio of superior grain weight, medium strength grain weight, inferior grain weight of rice contributed to the panicle weight under elevated [CO2] and warmed temperature |
2015年灌浆初期强势粒占穗质量比例高于2016年, 2015年强势粒占穗质量比例随水稻生育进程逐渐降低到收获期的9.9%~11.4%, 而2016年强势粒占穗质量比例降低速率较慢, 收获期降至14.7%~ 17.0%。2015年中势粒占穗质量比例随水稻生育期进程逐渐增加, 收获期为80.6%~84.8%, 2016年中势粒占穗质量比例生育期内变化不大, 收获期为73.2%~77.2%。2015年弱势粒占穗质量比例变化呈先增后减的趋势, 收获期为5.2%~7.6%, 2016年弱势粒占穗质量比例逐渐增加, 收获期为8.5%~10.6% (图 8)。
3 讨论本研究表明, [CO2]升高200 μmol·mol-1水稻产量增加了12.9%~14.8%, 温度每升高1 ℃产量减少8.6%~23.9%, 与前人研究的高[CO2]增加水稻产量和高温减少水稻产量的结果一致[7, 10, 19]。但[CO2]升高和温度升高相伴而生, 密不可分。尽管众多研究者通过试验或者模型等方式预测温度升高2 ℃以内, [CO2]升高可以弥补高温对作物产量的负效应[15]。但Wang等[6]与笔者于2015年和2016年在T-FACE平台上共同对南粳9108进行研究工作的结果表明, 高[CO2]和高温下水稻减产。Wang等[6]研究认为抽穗期单位面积干物质积累减少和孕穗期单茎氮素积累降低导致单位面积颖花密度(有效穗数×穗粒数)减少是减产的主要原因, 并将研究重点侧重于水稻齐穗前。灌浆期是紧跟着水稻齐穗期的产量形成的关键时期, 易受到[CO2]和温度等环境因素的影响。本研究侧重于不同粒位籽粒灌浆充实度对[CO2]和温度升高的响应。
水稻产量的构成因子主要为有效穗数、穗粒数、结实率和饱粒千粒质量。本试验平台研究表明, 高[CO2]增加了水稻有效穗数和减少了穗粒数, 增温则减少了有效穗数和穗粒数, 并且[CO2]升高对有效穗数和穗粒数的影响幅度高于增温。[CO2]和温度升高对有效穗数和穗粒数交互作用明显, 减少了有效穗和穗粒数。[CO2]和温度升高单位面积颖花密度是影响产量的主要因素, 两者的相关系数r2为0.73[6]。[CO2]和温度升高对结实率和饱粒千粒质量影响不明显。根据水稻粒位分布和饱粒的筛选方法, 结实率是饱粒数与(饱粒数+半饱粒数+瘪粒数)的比值, 饱粒包括强势粒和部分中势粒, 半饱粒是中势粒的一部分, 瘪粒可能是中、弱势粒的一部分。因此, 在[CO2]和温度升高下, 产量构成因子的结实率和饱粒千粒质量, 不能反映强、中、弱势粒对产量的影响程度。
本研究发现, 在各处理下, 不同粒位籽粒千粒质量从高到低表现为强、中、弱势粒。高[CO2]可以一定程度增加弱势粒千粒质量, 增温明显降低了中势粒千粒质量(表 3), [CO2]和温度升高中势粒千粒质量仍有所减少。[CO2]升高促进了水稻灌浆过程强、弱势穗粒质量增加, 降低了中势粒穗粒质量, 最终提高了水稻产量。高[CO2]和高温下中势粒质量及其占穗质量比例两年平均分别降低了10.8%和4.5%, 弱势粒占穗质量比例增加了33.1%, 中势粒、弱势粒占穗质量比例处于此消彼长的状态。就单穗水稻而言, 弱势粒占穗质量比例约10%, 而中势粒占穗质量比例在70%~ 85%之间, 中势粒千粒质量及其粒质量和比例远大于弱势粒, 中势粒的变化对水稻的影响程度远大于弱势粒。但高[CO2]和高温下大幅增加的弱势粒占穗质量比例降低了水稻充实度。因此, [CO2]和温度升高下, 弱势粒占穗质量比例的增加, 中势粒千粒质量、穗粒质量及其占穗质量比例的降低是水稻减产的重要原因。
水稻结实率具有不稳定性, 即在不同年度间、或同一年度不同地区间结实率忽高忽低, 甚至大起大落, 主要由于弱势粒结实(充实)不稳定, 强势粒在地区和年度间的变异相对较小[14, 20]。尽管[CO2]升高, 弱势粒灌浆速率增大, 粒质量提高, 但研究表明限制弱势粒质量的因素不是“源”, 而是弱势粒库容较小, 其着生的二次枝梗维管束导管面积和韧皮部面积较小, 导管、筛管和细胞分化数量较少, 进而造成无机物和有机物运输的“流”不畅[21]。弱势粒对环境和栽培条件反应具有敏感性, 同时弱势粒结实率和粒质量具有可调性[14]。因此, 通过改变水稻环境和栽培条件, 扩“源”疏“流”弱势粒, 稳定强势粒穗粒质量及其比例, 提高中势粒穗粒质量及其比例, 是缓解[CO2]和温度升高对水稻产量作用的重要途径, 尤其在相对高温多雨的年份。
相对2015年水稻季正常的温度和降雨分配, 2016年水稻季拔节期和开花期高温缺水、灌浆期持续阴雨, 导致高[CO2]和高温下2016年水稻产量明显低于2015年。并且, [CO2]升高下, 温度对水稻产量的负效应明显增加, 与Wang等[22]研究结果一致。究其原因, 第一, [CO2]持续升高造成全球尤其北半球高温、暴雨等灾害性天气频发[18, 23], 2016年灌浆期的长期阴雨天气显著降低了高[CO2]和高温下水稻光合作用[24], 减少了光合物质的积累;第二, 高[CO2]和高温加速了水稻的生育进程, 使水稻提前开花2~5 d, 生育期缩短3~10 d[6, 18, 25], 缩短了物质的积累时间, 阻碍了物质的积累, 而[CO2]升高导致气孔关闭, 蒸腾作用减弱, 热耗散降低[4, 7], 又加剧了增温的作用[22]。同时, 2016年水稻单穗质量和中势粒穗粒质量均显著低于2015年, 其中, 中势粒占穗质量比例比2015年约减少了10个百分点, 而弱势粒占穗质量比例约增加了3个百分点。但弱势粒千粒质量远低于强势粒和中势粒千粒质量。因此, 在相对高温多雨的稻季, [CO2]和温度升高下, 中势粒占穗质量比例下降和弱势粒占穗质量比例增加是水稻减产的重要原因。
4 结论(1) 高[CO2]增加了水稻产量, 高温减少了水稻产量。[CO2]升高不能弥补高温对水稻产量的负效应, 这与不同粒位籽粒的灌浆充实度有关。
(2) 水稻收获期强、中、弱势粒千粒质量依次降低, [CO2]和温度升高降低了单穗粒质量和中势粒千粒质量、穗粒质量及其占穗质量比例。由于中势粒占穗质量比例远高于强势粒和弱势粒的占比, 因此, 中势粒穗粒质量及其比例的减少对产量的影响占主导作用。
(3) [CO2]和温度升高增加了弱势粒占穗粒质量比例、减少了中势粒千粒质量和穗粒质量及比例, 降低了籽粒的充实度, 造成了水稻减产。
(4) 2016年单穗粒质量比2015年明显减小, 产量下降幅度大(17.3%~28.6%), 与2016年水稻开花期高温、灌浆期多雨有关。
| [1] |
Van Groenigen K J, van Kessel C, Hungate B A. Increased greenhousegas intensity of rice production under future atmospheric conditions[J]. Nature Climate Change, 2013, 3: 228-291. DOI:10.1038/nclimate1689 |
| [2] |
Kazemi S, Eshghizadeh H R, Zahedi M. Responses of four rice varieties to elevated CO2 and different salinity levels[J]. Rice Science, 2018, 25: 142-151. DOI:10.1016/j.rsci.2018.04.002 |
| [3] |
IPCC(The International Panel on Climate Change). Climate change 2013: The physical science basis[M]. Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge, UK, New York, USA: Cambridge University Press, 2013: 1-8.
|
| [4] |
Cai C, Yin X Y, He S Q, et al. Responses of wheat and rice to factorial combinations of ambient and elevated CO2 and elevated temperature in FACE experiment[J]. Global Change Biology, 2016, 22(2): 856-874. DOI:10.1111/gcb.13065 |
| [5] |
Cai C, Li G, Yang H L, et al. Do all leaf photosynthesis parameters of rice acclimate to elevated CO2, elevated temperature, and their combination, in FACE environments?[J]. Global Change Biology, 2018, 24(4): 1685-1707. DOI:10.1111/gcb.13961 |
| [6] |
Wang W L, Cai C, Lam S K, et al. Elevated CO2 cannot compensate for japonica grain yield losses under increasing air temperature because of the decrease in spikelet density[J]. European Journal of Agronomy, 2018, 99: 21-29. DOI:10.1016/j.eja.2018.06.005 |
| [7] |
Zhu C W, Zhu J G, Cao J, et al. Biochemical and molecular characteristics of leaf photosynthesis and relative seed yield of two contrasting rice cultivars in response to elevated[CO2][J]. Journal of Experimental Botany, 2014, 65(20): 6049-6056. DOI:10.1093/jxb/eru344 |
| [8] |
Matsui T, Omasa K, Horie T. High temperature at flowering inhibits swelling of pollen grains, a driving force for thecae dehiscence in rice (Oryza sativa L.)[J]. Plant Production Science, 2000, 3(4): 430-434. DOI:10.1626/pps.3.430 |
| [9] |
Madan P, Jagadish S V K, Craufurd P Q, et al. Effect of elevated CO2 and high temperature on seed-set and grain quality of rice[J]. Journal of Experimental Botany, 2012, 63(10): 3843-3852. DOI:10.1093/jxb/ers077 |
| [10] |
Xiong D L, Ling X X, Huang J L, et al. Meta-analysis and dose-response analysis of high temperature effects on rice yield and quality[J]. Environmental and Experimental Botany, 2017, 141: 1-9. DOI:10.1016/j.envexpbot.2017.06.007 |
| [11] |
万运帆, 游松材, 李玉娥, 等. CO2浓度和温度升高对早稻生长及产量的影响[J]. 农业环境科学学报, 2014, 33(9): 1693-1698. WAN Yun-fan, YOU Song-cai, LI Yu-e, et al. Influence of elevated atmospheric CO2 concentration and temperature on growth and yield of early rice[J]. Journal of Agro-Environment Science, 2014, 33(9): 1693-1698. |
| [12] |
Mohapatra P K. The time of flowering affects grain quality and spikelet partitioning within the rice panicle[J]. Australian Journal of Plant Physiology, 1993, 20: 231-240. |
| [13] |
盛婧, 陶红娟, 陈留根. 灌浆期不同时段温度对水稻结实与稻米品质的影响[J]. 中国水稻科学, 2007, 21(4): 396-402. SHENG Jing, TAO Hong-juan, CHEN Liu-gen. Response of seed setting and grain quality of rice to temperature at different time during grain filling period[J]. Chinese Journal Rice Science, 2007, 21(4): 396-402. DOI:10.3321/j.issn:1001-7216.2007.04.012 |
| [14] |
杨建昌. 水稻弱势粒灌浆机理与调控途径[J]. 作物学报, 2010, 36(11): 2011-2019. YANG Jian-chang. Mechanism and regulation in the filling of inferior spikelets of rice[J]. Acta Agronomica Sinica, 2010, 36(11): 2011-2019. |
| [15] |
Krishnan K, Swain D K, Bhaskar B C, et al. Impact of elevated CO2 and temperature on rice yield and methods of adaptation as evaluated by crop simulation studies[J]. Agriculture, Ecosystems and Environment, 2007, 122: 233-242. DOI:10.1016/j.agee.2007.01.019 |
| [16] |
刘钢, 韩勇, 朱建国, 等. 稻麦轮作FACE系统平台Ⅰ.系统结构与控制[J]. 应用生态报, 2002, 13(10): 1253-1258. LIU Gang, HAN Yong, ZHU Jian-guo, et al. Rice-wheat rotational FACE platformⅠ. System structure and control[J]. Chinese Journal of Applied Ecology, 2002, 13(10): 1253-1258. |
| [17] |
李杰, 张洪程, 龚金龙, 等. 不同种植方式对超级稻籽粒灌浆特性的影响[J]. 作物学报, 2011, 37(9): 1631-1641. LI Jie, ZHANG Hong-cheng, GONG Jin-long, et al. Influence of planting methods on grain-filling properties of super rice[J]. Acta Agronomica Sinica, 2011, 37(9): 1631-1641. |
| [18] |
刘建丰, 康春林, 伏军, 等. 水稻籽粒充实状况指标测定方法研究[J]. 作物研究, 1993, 7(1): 16-19. LIU Jian-feng, KANG Chun-lin, FU Jun, et al. Study on the method of determining indicators of rice grain filling[J]. Crop Research, 1993, 7(1): 16-19. |
| [19] |
凌霄霞, 张作林, 翟景秋, 等. 气候变化对中国水稻生产的影响研究进展[J]. 作物学报, 2019, 45(3): 323-334. LING Xiao-xia, ZHANG Zuo-lin, ZHAI Jing-qiu, et al. A review for impacts of climate change on rice production in China[J]. Acta Agronomica Sinica, 2019, 45(3): 323-334. |
| [20] |
Yang J C, Zhang J H. Grain filling problem in "super" rice[J]. Journal of Experimental Botany, 2010, 61(1): 1-5. DOI:10.1093/jxb/erp348 |
| [21] |
黄升谋, 邹应斌, 刘春林. 杂交水稻两优培九强、弱势粒结实生理研究[J]. 作物学报, 2005, 31(1): 102-107. HUANG Sheng-mou, ZOU Ying-bin, LIU Chun-lin. Setting physiology of the superior and inferior grains of hybrid rice Liangyoupeijiu[J]. Acta Agronomica Sinica, 2005, 31(1): 102-107. |
| [22] |
Wang D R, Bunce J A, Tomecek M B, et al. Evidence for divergence of response in Indica, Japonica, and wild rice to high CO2×temperature interaction[J]. Global Change Biology, 2016, 22: 2620-2632. DOI:10.1111/gcb.13279 |
| [23] |
Millar R J, Fuglestvedt J S, Friedlingstein P, et al. Emission budgets and pathways consistent with limiting warming to 1.5℃[J]. Nature Geoscience, 2018, 11(6): 454-455. DOI:10.1038/s41561-018-0153-1 |
| [24] |
袁嫚嫚, 朱建国, 刘钢, 等. 不同天气水稻光合日变化对大气CO2浓度和温度升高的响应——FACE研究[J]. 生态学报, 2018, 38(6): 1899-1907. YUAN Man-man, ZHU Jian-guo, LIU Gang, et al. Response of diurnal variation of photosynthesis to elevated atmospheric CO2 concentration and temperature of rice between cloudy and sunny day:A FACE study[J]. Acta Ecologica Sinica, 2018, 38(6): 1899-1907. |
| [25] |
景立权, 赖上坤, 王云霞, 等. 大气CO2浓度和温度互作对水稻生长发育的影响[J]. 生态学报, 2016, 36(14): 4254-4265. JING Li-quan, LAI Shang-kun, WANG Yun-xia, et al. Combined effect of increasing atmospheric CO2 concentration and temperature on growth and development of rice:A research review[J]. Acta Ecologica Sinica, 2016, 36(14): 4254-4265. |