 2021, Vol. 40
2021, Vol. 40



2. 南京信息工程大学江苏省农业气象重点实验室, 南京 210044
2. Jiangsu Key Laboratory of Agricultural Meteorology, Nanjing University of Information Science and Technology, Nanjing 210044, China
IPCC第五次评估报告指出,近130年来人为活动导致的全球地表平均温度升幅已达0.85 ℃[1],预计2030—2052年将达1.5 ℃[2]。气候变暖存在季节和昼夜非对称性[3]。近50年来江淮地区年平均气温增温速率为0.028 ℃·a-1,冬春季增温趋势明显高于夏秋季,全年最低气温增幅高于最高气温,夜间气温增幅大于日间气温[4-5]。随气温增加,平均地温也呈明显升高趋势[6]。
温度是影响水稻生长发育最重要的环境要素之一。光合作用是作物生产和能量代谢的基础生理过程,极易受温度变化影响[7]。通过快速叶绿素荧光技术可实现无损条件下对植物光合性能的活体监测,利用高速连续激发光得到的叶绿素快相荧光动力学曲线(OJIP曲线),可全面展现位于类囊体膜上的光系统Ⅱ(PSⅡ)的光反应过程、效率和结果[8]。研究表明,夜间增温抑制水稻光合特性,降低蒸腾速率(Tr)和气孔导度(Gs)以调节水分散失和能量循环,显著降低叶片净光合速率(Pn)和胞间CO2浓度(Ci)[9-10],但也有报道认为夜间增温可缓解低温对叶片抑制性以增加Pn[11]。夜间增温使灌浆期水稻荧光耗散增多,性能指数(PIABS、PICS和PITotal)降低,光合性能下降,但较少研究涉及水稻OJIP曲线[12]。
长江中下游地区是我国水稻种植的主要区域。水稻是典型的喜硅作物,对硅的吸收存在主动过程,其穗中硅含量约为10%~15%[13],远高于氮磷钾含量。施用硅肥可有效增加水稻的Pn和Gs,提高干物质积累和产量,能缓解夜间增温引起的抑制效应[14-16]。生物炭富含有机碳,可明显降低土壤容重,增加土壤含水量,改善土壤结构[17]。生物炭可提高水稻叶绿素含量和持绿时间,增强光合能力[18-19]。在增温条件下,生物炭能有效增强土壤酶活性[20],与矿质肥料混用可促进水稻生长,提高产量和品质[21]。
夜间增温、施用生物炭或硅肥单因子或两因子对水稻生产的影响已有研究,但三者耦合对水稻光合及荧光特性有何影响,尚缺少相关报道。本文通过大田模拟试验,探讨施生物炭和硅肥能否缓解夜间增温对水稻光合和荧光特性的不利影响,为后续研究气候变暖背景下水稻生产调控和高产优质栽培提供试验依据。
1 材料与方法 1.1 试验区概况田间试验在南京信息工程大学农业气象试验站(32.0°N,118.8°E)进行,时间为2019年6月至10月。该站地处亚热带湿润气候区,年均降水量1 100 mm,年均气温15.6 ℃,2019年水稻生长季平均气温为26.52 ℃。供试土壤为潴育型水稻土,灰马肝土属,质地为壤质黏土,全碳、全氮和黏粒含量分别为19.4、1.45 g·kg−1和26.1 g·kg−1,pH 6.2(1∶1土水比)。供试生物炭为稻壳生物炭,由天津亚德尔生物质科技股份有限公司提供,含碳量约50%。供试硅肥为钢渣粉和矿粉。供试生物炭和硅肥的基本理化性质见表 1。供试水稻为南粳5055,该品种株高适中,抗倒伏性强,适宜在江苏沿江及苏南地区种植,2019年5月10日育苗,6月14日移栽。
|
|
表 1 供试生物炭及硅肥基本性质 Table 1 Basic properties of tested biochar and silicon fertilizer |

本试验采用3因素3水平正交试验设计,3因素为夜间增温、施生物炭和施硅,每因素设3水平,共9个处理。夜间增温设3水平,即常温对照(W0)、5 mm膜覆盖(W1)和11 mm膜覆盖(W2);生物炭3水平,为不施生物炭(B0)、施7.5 t·hm-2生物炭(B1)和施17.5 t·hm-2生物炭(B2);施硅3水平,为不施硅(Si0)、施钢渣粉(Si1,200 kg·hm-2)和施矿粉(Si2,200 kg·hm-2)。
夜间增温采用开放式被动增温方法,自2019年6月14日至10月14日,夜间(19:00—6:00)用铝箔反光膜覆盖稻麦冠层,根据水稻生长进程对铝箔膜的高度进行调整,使铝箔膜与水稻冠层保持0.3 m左右。为避免铝箔膜覆盖对降雨的阻挡和大风对增温设施的破坏(风速>10 m·s-1),雨天和大风天气下夜间不进行覆盖。采用温度自动记录仪记录水稻田5 cm土层和10 cm土层温度。生物炭和硅肥均在2019年6月13日翻耕农田时作为基肥施入土壤,同时各小区施用复合肥315 g(15∶15∶15)。小区面积为2 m×2 m=4 m2,随机排列共9个小区。除7月28日到8月12日(晒田期)外,田间水位保持5 cm左右。
1.3 测定方法水稻移栽后,每周用Li-6400便携式光合作用仪,测定叶片Pn、Ci、Tr和Gs等光合和蒸腾参数,测定前预热0.5 h,测定时间为8:00—11:00,每小区重复3次取平均值。
2019年9月12日上午8:00—11:00(灌浆期),用植物效率分析仪(Pocket PEA)测定叶片荧光动力学曲线(OJIP曲线)和荧光参数,每小区选有代表性的3株水稻,用暗适应叶夹使水稻叶片处于暗反应状态20 min后再进行测定,3次数据取平均值。
2019年10月15日(成熟期)进行田间考种,在小区中心区域选取考种样区(0.5 m×0.5 m=0.25 m2),剪下稻穗后用脱谷机处理,称量质量记录各小区产量。
1.4 数据处理叶绿素快相荧光动力曲线(OJIP曲线)是以时间为横坐标、原始荧光值为纵坐标的曲线,可根据公式ΔVt=(Ft-Fo)(/ Fm-Fo)对原始曲线进行标准化处理,使各处理更具可比性,体现各特征位点的微小差异。
气孔限制值(Ls)可用公式Ls=1-Ca/Ci进行计算,其中Ca和Ci分别为大气和胞间CO2浓度,μmol · mol-1。
水分利用效率(WUE)计算公式则为WUE=Pn/ Tr,即净光合速率与蒸腾速率之比。
采用Microsoft Excel 2007进行数据整理汇总和极差分析,利用SPSS 19.0进行单因素方差分析(Oneway ANOVA),采用Origin 9.1进行图表绘制。
2 结果与分析 2.1 被动式增温效果水稻全生育期夜间平均土温变化见图 1。在夜间时段(19:00—6:00),除W0处理的5 cm土层温度在6:00略有回升,其余增温处理下土温均表现为降低趋势。其中5 cm土层温度变化幅度大于10 cm土层,说明淹水灌溉下稻田水层对土温变化有一定的缓冲作用,土温对环境温度变化的响应会随土层深度的增加而变得迟钝。

|
图 1 水稻全生育期夜间平均土壤温度变化 Figure 1 Change of average soil temperature at night during the whole growth period of rice |
相比于W0处理,W1和W2处理下的5 cm土层夜间均温分别增加了0.72 ℃和0.28 ℃,而10 cm土层温度增幅则是0.43 ℃和0.05 ℃,说明铝箔膜的覆盖对土壤有一定的增温效应。
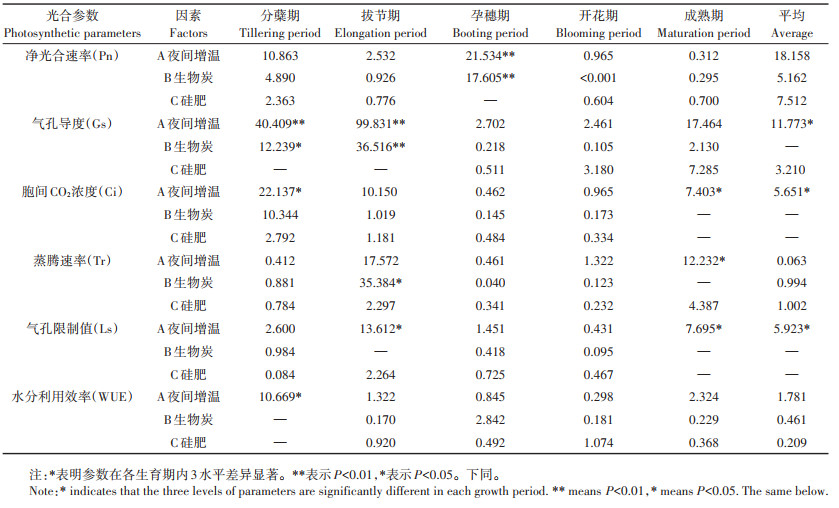
2.2 光合参数 2.2.1 净光合速率(Pn)各处理下Pn全生育期变化趋势均表现为先升后降(图 2A)。其中,W0和W1多在孕穗期达最大值,W2均在开花期,且各处理间拔节、孕穗和开花期Pn差异明显,说明膜厚度增加会使水稻Pn达到最大值的时期有所延后,水稻Pn在关键生育期受不同田间处理的影响最大。

|
不同字母表示同一生育期不同处理间的差异显著(P < 0.05) Different letters indicated significant differences among different treatments in the same growth period(P < 0.05) 图 2 不同处理对水稻各生育期光合参数的影响 Figure 2 Effects of different treatments on photosynthetic parameters of rice in each growth period |
各处理全生育期Pn平均值中W2B2Si1均值最低,W1B1Si2均值最高。由表 2可知,夜间增温和生物炭均在水稻孕穗期达极显著水平(P < 0.01),但3因素对全生育期平均Pn的影响均未达显著(P>0.05)。
|
|
表 2 不同处理下水稻各生育期光合参数的F值 Table 2 F value of photosynthetic parameters of rice in each growth period under different treatments |
相同夜间增温处理下水稻Gs随生育期推进变化趋势相似,且W2处理下Gs在分蘖、拔节和孕穗期基本低于其他处理,而生育期后期相对较高(图 2B)。其中W0和W1基本在拔节期达到最大值(除W0B1Si1和W1B1Si2),W2则均在开花期;各处理下Gs最小值均在成熟期。
对全生育期Gs平均值分析可知,W2B2Si1均值最低,W1B2Si0均值最高。根据方差分析(表 2),夜间增温在分蘖和拔节期影响极显著(P < 0.01),生物炭则在分蘖和拔节期分别达到显著(P < 0.05)和极显著水平(P < 0.01),且夜间增温可显著影响全生育期平均Gs(P < 0.05)。
2.2.3 胞间CO2浓度(Ci)各处理下Ci的变化趋势基本表现为先升后降(图 2C),除W0B2Si2和W1B0Si1外,W0和W1处理下Ci均在开花期达到最大值,W2则均在孕穗期。相比于W0处理下Ci均在成熟期达到最小值,W1和W2的Ci最小值有所提前,大多在分蘖期。
全生育期Ci平均值中,W2B2Si1均值最低,W1B2Si0均值最高。进行方差分析可知(表 2),夜间增温下水稻Ci在分蘖和成熟期差异显著(P < 0.05),并显著影响全生育期平均Ci(P < 0.05)。
2.2.4 蒸腾速率(Tr)不同田间处理下的Tr最大值基本处于生育期前期(分蘖、拔节和孕穗期),最小值则均出现在成熟期(图 2D)。Tr在孕穗和开花期的变化趋势与Gs不一致,可能与夏季高温、饱和水汽压差及所处生殖阶段等有关。
全生育期Tr平均值中W0B1Si1均值最低,W1B2Si0均值最高。由表 2可知,夜间增温下水稻Tr在成熟期均差异显著(P < 0.05),生物炭在拔节期达显著水平(P < 0.05),但3因素对全生育期平均Tr均无显著影响(P>0.05)。
2.2.5 气孔限制值(Ls)Ls在不同的增温处理下有不同的变化趋势,W0和W2基本表现为先升后降,并在成熟期有明显回升;W1则为“W”型,在拔节和开花期达到较小值(图 2E)。除W2B1Si0外,各处理下Ls均在成熟期达到最大值,而W2处理下最小值均在分蘖期,其余处理多在开花期。
全生育期Ls平均值中,W1B2Si0均值最低,W0B2Si2均值最高。由方差分析表可知(表 2),夜间增温在拔节和成熟期均差异显著(P < 0.05),对全生育期平均Ls的影响也达显著水平(P < 0.05)。
2.2.6 水分利用效率(WUE)各处理下WUE多在分蘖到孕穗期表现为上升趋势,孕穗到成熟期为先降后升;而W0B2Si2、W2B0Si2和W2B2Si1均呈现上升趋势,W1B0Si1则在拔节和开花期达到较小值(图 2F)。WUE的最大值均出现在成熟期,最小值除W1B0Si1和W1B2Si0外均出现在分蘖期。
根据全生育期WUE平均值可知W2B0Si2均值最低,W0B0Si0均值最高。由表 2可知,夜间增温下水稻WUE在分蘖期差异显著(P < 0.05),其余时期均无显著影响(P>0.05)。
2.2.7 光合参数的极差分析由极差分析图(图 3)可知,3因素对各光合参数的影响程度不同,其中Pn、Gs和Ls均为夜间增温>硅肥>生物炭,Ci和WUE为夜间增温>生物炭>硅肥,而Tr则为硅肥>生物炭>夜间增温,说明夜间增温对水稻光合作用影响最大。其中,5 mm铝箔膜覆盖可增强Gs和Ci,但Ls会明显降低;随膜厚度的增加,夜间增温对Pn和WUE的抑制作用更为明显,但对Tr有促进作用。对于生物炭,施入7.5 t·hm-2会抑制Gs和Tr,促进Pn、Ci和WUE,而17.5 t·hm-2处理下对这些参数作用效果相反;不同施炭量均能有效促进Ls。相比于不施硅肥,矿粉能有效促进Gs和Tr,但钢渣粉则有抑制作用,这与对Ls的影响相反;不同种类的硅肥均能有效促进Pn和WUE,抑制Ci,其中矿粉对Pn的促进作用更明显。

|
图 3 不同处理对水稻全生育期光合参数平均值的极差分析 Figure 3 Range analysist of different treatments on the average photosynthetic parameters of rice during the whole growth period |
Pn、Gs、Ci、Tr、Ls和WUE达到最高的最佳因素组合分别为W0B1Si2、W1B2Si2、W1B1Si0、W1B2Si2、W0B2Si1和W0B1Si1。依据综合平衡法,参考光合参数的最佳组合和3因素影响程度可以选出最适组合为W0B2Si2,即夜间不覆盖铝箔膜、施加17.5 t·hm-2的生物炭和200 kg·hm-2的矿粉。
2.3 荧光特性各处理在灌浆期均有典型的叶绿素快相荧光动力学曲线,曲线形态较为类似,均拥有O、K、J、I和P相点(图 4A)。各处理到达P相点的时间略有不同,W0B1Si1、W1B2Si0、W2B0Si2和W2B2Si1均为0.29 s,而W2B1Si0为0.3 s,其余处理则均为0.4 s。夜间增温、施加生物炭和钢渣粉使水稻到达P相点时间提前,而矿粉则有推迟效果。由标准化荧光差值图(图 4B)可知,灌浆期的标准化荧光值差异主要集中于K相和I相之间,除W0B0Si0和W0B2Si2在I相外,其余处理均在J相附近差异最大,这表明各处理会不同程度地影响PSⅡ反应中心QA-往QB的电子传递。从整体上看,在O - P相之间,除I相处个别处理有高于W0B0Si0的窄峰,其余时刻基本上是W0B0Si0的标准化荧光值最高,W0B2Si2的最低。

|
图 4 灌浆期叶绿素快相荧光动力学曲线 Figure 4 Chlorophyll fast phase fluorescence kinetics curve during filling period |
由雷达图(图 5)可知,各处理间Vj差异较大,W0B2Si2处理下Vj明显低于其他处理,这表明2 ms时该处理下水稻叶片活性反应中心关闭程度低,可明显促进PSⅡ受体测的电子传递。Vi差异不明显,其中W0B0Si0处理下Vi最低,方差分析(表 3)可知3因素对Vi均有显著影响(P < 0.05)。

|
Vj为J相的相对可变荧光强度;Vi为I相的相对可变荧光强度;Fo/ Fm为热耗散量子比率;Fv/Fm为PSⅡ反应中心捕获的最大量子产额;Fv/ Fo为PSⅡ潜在活性;ABS/RC为单位活性反应中心吸收光量;DIO/RC为单位活性反应中心热耗散的能量;TRO/RC为单位活性反应中心捕获的光量;ETO/RC为单位活性反应中心用于电子传递的能量;ABS/CSO为单位激发态面积吸收的光量;DIO/CSO为单位激发态面积热耗散的能量;TRO/CSO为单位激发态面积被反应中心捕获的光量;ETO/CSO为单位激发态面积用于电子传递的能量;PIABS为以吸收光能为基础的性能参数;PICSo为以吸收基础荧光(Fo)为基础的性能指数;PICSm为以吸收最大荧光(Fm)为基础的性能指数;DFABS为以吸收光能为基础的推动力指数;DFTotal为综合叶片推动力指数。下同 Vj is the relative variable fluorescence intensity at the J-step; Vi is the relative variable fluorescence intensity of phase I; Fo / Fm is the heat dissipation quantum ratio; Fv/Fm is the maximum quantum yields captured by PSⅡreaction center; Fv/Fo is the PSⅡpotential activity; ABS/RC is the absorption flux per RC(RC is per QA-reducing PS Ⅱ reaction center); DIO/RC is the dissipated energy flux per RC; TRO/RC is the trapped energy flux per RC; ETO/RC is the electron transport flux per RC; ABS/CSO is the absorption flux per CS(CS is per excited cross section); DIO /CSO is the dissipated energy flux per CS; TRO/CSO is the trapped energy flux per CS; ETO / CSO is the electron transport flux per CS; PIABS is the performance index on absorption basis; PICSo is the performance index on cross section basis(at t=0);PICSm is the performance index on cross section basis(at t= tm); DFABS is the driving force index based on absorbing light energy; DFTotal is the comprehensive blade driving force index. The same below 图 5 灌浆期叶绿素荧光参数比较雷达图 Figure 5 Comparison of chlorophyll fluorescence parameters during the filling period |
|
|
表 3 不同处理下水稻灌浆期荧光参数的F值 Table 3 F value of fluorescence parameters of rice at the filling period under different treatments |
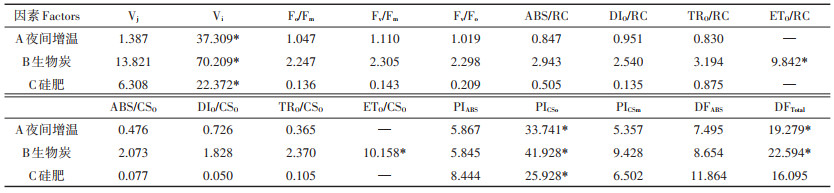
比活性参数,即表示活跃的单位反应中心或单位受光面积的各种量子效率(ABS/RC、DIO/RC、TRO/RC、ETO/RC、ABS/CSO、DIO/CSO、TRO/CSO和ETO/CSO)。由雷达图可知(图 5),比活性参数差异较小,W2B2Si1处理下各参数均高于其他处理,说明该处理下PSⅡ活动中心的光化学活性最强。对比活性参数进行方差分析(表 3),ETO/RC和ETO/CSO受生物炭的影响均达显著水平(P < 0.05)。
相比于Fv/Fm,性能指数(PIABS、PICSo和PICSm)和驱动力(DFABS和DFTotal)更能准确地反映植物光合机构的状态,体现胁迫对光合机构的影响。由雷达图(图 5)可知,W2B1Si0处理下性能参数和驱动力最低,而W0B2Si2则明显高于其他处理,这说明夜间增温对水稻光合性能有一定抑制作用,而生物炭和硅肥则可加以改善。根据方差分析表(表 3)可知,3因素对PICSo的影响均达显著水平(P < 0.05),而DFTotal受夜间增温和生物炭的影响显著(P < 0.05)。
由极差图(图 6)分析可知,夜间增温可以提高I相和J相处的相对可变荧光强度,生物炭对Vj的抑制作用随施用量增加而增强,但对Vi作用效果相反,而两种硅肥对两者均有抑制作用。在这些参数中,Fo/ Fm、DIO/RC和DIO/CSO可以反映光合器官的热耗散,夜间增温和生物炭均有明显的促进作用,且会随施炭量增加而增强,硅肥则能降低植株热耗散,3因素对热耗散参数的影响程度均为生物炭>夜间增温>硅肥。能反映PSⅡ活动中心的光化学活性的参数有Fv/Fm、Fv/Fo和其他比活性参数,夜间增温和生物炭能有效抑制Fv/Fm和Fv/Fo;而其他比活性参数基本均会受夜间增温和生物炭的影响而升高,硅肥(除ETO/RC和ETO/ CSO)则多为抑制作用,且3因素对这些参数的影响程度均为生物炭>夜间增温>硅肥。夜间增温和7.5 t· hm-2生物炭会明显降低植株的光合性能指数和驱动力,而17.5 t·hm-2生物炭和硅肥则有明显的促进作用,且生物炭和硅肥的影响程度基本高于夜间增温。依据综合平衡法可知,使水稻灌浆期光合作用最强的组合为W0B2Si2,即夜间不覆盖铝箔膜、施加17.5 t· hm-2的生物炭和200 kg·hm-2的矿粉。

|
图 6 不同处理对灌浆期叶绿素荧光参数的极差分析 Figure 6 Range analysis of chlorophyll fluorescence parameters of different treatments at the filling period |
根据极差分析可知(图 7),3因素对水稻产量影响程度由大到小依次为夜间增温、硅肥和生物炭,其中W0B1Si0产量最高,为11.73 t·hm-2,W2B0Si2产量最低,为7.47 t·hm-2。夜间增温和硅肥均会降低水稻产量,W1和W2处理下产量较W0分别降低了27.49% 和20.11%,而Si1和Si2会使水稻减产15.13% 和18.94%;生物炭则有明显的增产效果,B1和B2较B0分别增产15.84%和10.94%。

|
图 7 不同处理对水稻产量的极差分析 Figure 7 Range analysis of rice yield under different treatments |
温度是影响水稻光合作用最敏感的环境因子之一,5 mm和11 mm铝箔膜可分别使5 cm土层夜间均温增加0.72 ℃和0.28 ℃,10 cm土层增加0.43 ℃和0.05 ℃。W2处理下各层土壤均温低于W1,原因可能是观测小区为W1B2Si0和W2B1Si0,施炭量的增加会加深土壤颜色,促使土层吸收较多的夜间水面辐射而进一步增加土温[22]。而在W0处理下,10 cm土层夜间均温变化幅度更大,这可能是由于施用生物炭会增加土壤有机质含量进而降低土壤热导率[23],使热量往深层土壤传递变慢,减缓W1和W2处理下的温度变化幅度。
光合参数可反映水稻在不同处理下光合作用的变化。本试验中,夜间增温降低水稻全生育期平均Pn、Ls和WUE,提高Tr、Ci和Gs,但11 mm铝箔膜覆盖会使Ci和Gs降低。高温会通过气孔限制和非气孔限制因素影响作物Pn,Ci降低和Ls升高表明气孔限制是主要原因,而Ci升高和Ls降低则认为是非气孔限制因素所致[24-25]。5 mm铝箔膜覆盖下Ls降低,Ci升高,Pn变化趋势与Ls一致,这可能是水稻光合器官损坏、细胞内光合酶活力降低等非气孔限制原因导致[25],与他人研究结果一致[14, 26]。11 mm铝箔膜覆盖下,Pn、Gs和Ci均表现为降低趋势,原因可能是Gs下降阻碍CO2进入细胞,水稻植株体内CO2供应不足,进而降低Pn,抑制光合作用[27];而Gs降低的同时Tr升高,这可能是高温导致叶室温度增加,叶片内外蒸汽压差随之增加导致的[28]。
施7.5 t·hm-2生物炭能促进Pn和Ci,这可能是生物炭通过增加通气孔隙、改善养分供应,促进水稻根系生长,保持叶片良好的功能状态[19, 29];但17.5 t·hm-2处理下抑制Pn和Ci,这可能是Ls升高导致气孔限制因素占主导作用[27]。两种硅肥均能提高Pn,降低Ci,可能是硅在叶表皮层沉积以增加叶片厚度和改善株型,提高上部高效叶面积以促进光合作用[30-31]。叶片中硅化细胞散射光透过量是绿色细胞的10倍,能促进水稻叶片对光能的吸收[32]。而Ci降低可能是光诱导过程中叶肉细胞的光合活性随光强增强而增高导致[33]。因此,适量生物炭和硅肥可通过改善水稻生理性状,以提高光合性能,进而缓解夜间增温带来的不利影响。施7.5 t·hm-2生物炭可增加WUE和Ls,降低Gs和Tr;施17.5 t·hm-2生物炭更能促进Ls,而其余参数呈现相反变化趋势。光合参数在不同施炭量下变化不一样,这可能是由于施炭量不同会影响土壤养分组成和理化性质,进而影响水稻根系的养分吸收和地上部光合作用[29, 34]。施加钢渣粉会增加Ls和WUE,降低Gs和Tr;矿粉则会增加Gs、Tr和WUE,降低Ls。钢渣粉使Tr降低,可能是硅能使表皮细胞沉积形成“角质-硅双层”结构而减少角质层蒸腾[14-15],并增加木质部导管的亲水性,降低管内水分流速,减少蒸腾[35];矿粉处理下Tr有所升高,这可能是由于水稻Pn和Gs增加促进了水稻光合作用,进而增强水稻叶片蒸腾作用。
自然环境中叶绿素荧光和光合作用密切相关,能反映光合过程中光系统对光能的吸收、传递和耗散等方面,因此,参数变化更能阐明不同处理下光合作用“内在性”变化[36]。比活性参数中ABS/RC和TRO/RC能反映天线色素增减[37],天线色素吸收光能多被PSⅡ反应中心捕获,用于电子传递和CO2固定,少数以荧光和热能形式耗散[38]。本研究表明,夜间增温处理虽一定程度上增强灌浆期水稻叶片PSⅡ光化学活性,但也会促使天线色素增多,以增加光能热耗散,并显著降低光合性能和PSⅡ原初光能转化效率,与他人研究结果一致[7, 12],这可能是夜间增温下光合作用受到抑制的原因之一。生物炭对水稻叶片的影响与夜间增温相似,但17.5 t·hm-2处理下能明显增强水稻的光合性能,这可能是大量生物炭能促进光合机构中QA- 到QB的电子传递,增加叶绿素含量,进而提高水稻的光合速率。硅肥则会促进PSⅡ原初光能转化效率,降低热耗散和PSⅡ光化学活性,原因可能是天线色素降低,使单个反应中心光能吸收和能量捕获速率减缓,进而降低水稻叶片对光能的吸收[39]。施用17.5 t· hm-2生物炭和硅肥可明显增强水稻灌浆期的光合性能指数和驱动力,而两种硅肥中矿粉更具效果,且施用生物炭和硅肥的影响程度均高于夜间增温,说明施生物炭和硅肥可有效缓解夜间增温对水稻灌浆期光合作用的抑制。根据光合和荧光参数可知,使水稻光合作用最强的处理组合为W0B2Si2,即夜间不覆盖铝箔膜、施17.5 t·hm-2生物炭和200 kg·hm-2矿粉。
夜间增温会明显降低水稻产量,W1和W2处理较W0分别减产27.49% 和20.11%,这可能是夜间增温下水稻Pn降低,光能热耗散增强,进而减少籽粒中光合产物。不同种类的硅肥均会导致水稻减产,这可能是由于供试稻田多年施硅导致土壤中有效硅含量偏高,超出稻田施硅肥临界值,抑制产量增长[40]。生物炭则有明显的增产效果,B1和B2分别增产15.84% 和10.94%,这可能是由于生物炭通过固定土壤N素来供应水稻生长后期营养需求,延缓衰老进程,保证充足光合作用,从而增加产量[18-19]。
4 结论(1)5 mm和11 mm铝箔膜覆盖水稻冠层,可分别使5 cm土层夜间均温增加0.72 ℃和0.28 ℃,10 cm土层温度增加0.43 ℃和0.05 ℃,增温效果明显。
(2)夜间增温降低水稻全生育期平均Pn、Ls和WUE,提高Tr、Ci和Gs,但11 mm铝箔膜覆盖下Ci和Gs降低;同时增加荧光耗散,抑制PSⅡ原初光能转化效率和光合性能,其中对Ls、Gs、Ci和光合性能的影响均达显著水平(P < 0.05)。施7.5 t·hm-2生物炭和硅肥可提高水稻叶片Pn,改善作物生长发育,进而缓解夜间增温对水稻光合作用和荧光特性的抑制作用。
(3)根据经济效益和综合平衡法可知,在增温和不施硅条件下,施用生物炭(7.5 t·hm-2)在促进光合作用的同时实现水稻增产。未来可进一步探讨夜间增温下施生物炭和硅肥对水稻生长发育、产量和温室气体排放影响,提高水稻生产应对气候变化的能力,为区域水稻可持续生产提供试验依据。
| [1] |
IP CC. Climate change 2013:The physical science basis. Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Cambridge: Cambridge University Press, 2014.
|
| [2] |
IP CC. Special report on global warming of 1.5℃(SR15)[M]. Cambridge: Cambridge University Press, 2018: 785.
|
| [3] |
谭凯炎, 房世波, 任三学, 等. 非对称性增温对农业生态系统影响研究进展[J]. 应用气象学报, 2009, 20(5): 634-641. TAN Kai-yan, FANG Shi-bo, REN San-xue, et al. Asymmetric trends of daily maximum and minimum temperature in global warming and its effects on agriculture ecosystems[J]. Journal of Applied Meteorological Science, 2009, 20(5): 634-641. |
| [4] |
黄爱军, 陈长青, 类成霞, 等. 江淮地区农业气候资源演变特征及作物生产应对措施[J]. 南京农业大学学报, 2011, 34(5): 7-12. HUANG Ai-jun, CHEN Chang-qing, LEI Cheng-xia, et al. Change characteristics of agricultural climatic resources and adaptation measures for crop production in Jianghuai region[J]. Journal of Nanjing Agricultural University, 2011, 34(5): 7-12. |
| [5] |
钟晨晨, 崔宁博, 谭畅, 等. 近53 a长江流域气温的时空变化特征分析[J]. 灌溉排水学报, 2016, 35(12): 88-96. ZHONG Chen-chen, CUI Ning-bo, TAN Chang, et al. Spatiotemporal of Yangtze River in 53 years[J]. Journal of Irrigation and Drainage, 2016, 35(12): 88-96. |
| [6] |
林洁, 陈效民, 张勇. 太湖典型水稻种植区近50年来气候变化特征及其对浅层土壤温度的影响[J]. 土壤, 2012, 44(5): 853-861. LIN Jie, CHEN Xiao-min, ZHANG Yong. Characteristics of climate change and their influences on shallow soil temperature in typical paddy planting area of Tai Lake region during 1960-2009[J]. Soils, 2012, 44(5): 853-861. |
| [7] |
刘春溪, 孙备, 王国骄, 等. 开放式增温对粳稻光合作用和叶绿素荧光参数的影响[J]. 生态环境学报, 2018, 27(9): 1665-1672. LIU Chun-xi, SUN Bei, WANG Guo-jiao, et al. Effects of open temperature increasing on photosynthesis and chlorophyll fluorescence parameters of japonica rice[J]. Ecology and Environmental Sciences, 2018, 27(9): 1665-1672. |
| [8] |
孟鹏, 安宇宁, 白雪峰. 沙地赤松光合及叶绿素a快相荧光动力学特性[J]. 生态学报, 2016, 36(11): 3469-3478. MENG Peng, AN Yu-ning, BAI Xue-feng. Photosynthetic characteristics and chlorophyll a fluorescence induction parameters of Pinus densiflora on sandy soil[J]. Acta Ecologica Sinica, 2016, 36(11): 3469-3478. |
| [9] |
Rehmani M I A, Wei G B, Hussain N, et al. Yield and quality responses of two indica rice hybrids to post-anthesis asymmetric day and night open-field warming in lower reaches of Yangtze River delta[J]. Field Crops Research, 2014, 156(2): 231-241. |
| [10] |
Mohammed A R, Tarpley L. High nighttime temperatures affect rice productivity through altered pollen germination and spikelet fertility[J]. Agricultural and Forest Meteorology, 2008, 149(6): 999-1008. |
| [11] |
陈金, 田云录, 董文军, 等. 东北水稻生长发育和产量对夜间升温的响应[J]. 中国水稻科学, 2013, 27(1): 84-90. CHEN Jin, TIAN Yun-lu, DONG Wen-jun, et al. Responses of rice growth and grain yield to nighttime warming in northeast China[J]. Chinese Journal of Rice Science, 2013, 27(1): 84-90. |
| [12] |
李君, 娄运生, 马莉, 等. 夜间增温和水分管理耦合对水稻叶片光合作用和荧光特性的影响[J]. 江苏农业学报, 2019, 35(3): 506-513. LI Jun, LOU Yun-sheng, MA Li, et al. Effect of nighttime warming and water managements coupling on photosynthetic and fluorescence characteristics in rice[J]. Jiangsu Journal of Agricultural Sciences, 2019, 35(3): 506-513. |
| [13] |
宫海军, 陈坤明, 王锁民, 等. 植物硅营养的研究进展[J]. 西北植物学报, 2004, 24(12): 2385-2392. GONG Hai-jun, CHEN Kun-ming, WANG Suo-min, et al. Advances in silicon nutrition of plants[J]. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(12): 2385-2392. |
| [14] |
郑泽华, 娄运生, 左慧婷, 等. 施硅对夜间增温条件下水稻叶片生理特性的影响[J]. 中国农业气象, 2017, 38(10): 663-671. ZHENG Ze-hua, LOU Yun-sheng, ZUO Hui-ting, et al. Effect of silicate application on rice physiological properties under nighttime warming[J]. Chinese Journal of Agrometeorology, 2017, 38(10): 663-671. |
| [15] |
Kim S G, Kim K W, Park E W, et al. Silicon induced cellwall fortification of rice leaves: A possible cellular mechanism of enhanced host resistance to blast[J]. Phytopathology, 2002, 92(10): 1095-1103. |
| [16] |
郑泽华, 娄运生, 左慧婷, 等. 施硅对夜间增温条件下水稻生长和产量的影响[J]. 中国农业气象, 2018, 39(6): 390-397. ZHENG Ze-hua, LOU Yun-sheng, ZUO Hui-ting, et al. Effect of silicate application on hybrid rice growth and yield under nighttime warming[J]. Chinese Journal of Agrometeorology, 2018, 39(6): 390-397. |
| [17] |
王冲, 王玉峰, 谷学佳, 等. 连续施用生物炭对黑土基础理化性质的影响[J]. 土壤通报, 2018, 49(2): 428-434. WANG Chong, WANG Yu-feng, GU Xue-jia, et al. Effects of continuous application of biochar on the basic physicochemical properties of black soil[J]. Chinese Journal of Soil Science, 2018, 49(2): 428-434. |
| [18] |
王悦满, 高倩, 薛利红, 等. 生物炭不同施加方式对水稻生长及产量的影响[J]. 农业资源与环境学报, 2018, 35(1): 58-65. WANG Yue-man, GAO Qian, XUE Li-hong, et al. Effects of different biochar application patterns on rice growth and yield[J]. Journal of Agricultural Resources and Environment, 2018, 35(1): 58-65. |
| [19] |
陈盈, 张满利, 刘宪平, 等. 生物炭对水稻齐穗期叶绿素荧光参数及产量构成的影响[J]. 作物杂志, 2016, 32(3): 94-98. CHEN Ying, ZHANG Man-li, LIU Xian-ping, et al. Effects of biochar on chlorophyll fluorescence at full heading stage and yield components of rice[J]. Crops, 2016, 32(3): 94-98. |
| [20] |
蒋容, 余一, 唐玉蓉, 等. 增温和生物炭添加对农田土壤酶活性的影响[J]. 四川农业大学学报, 2018, 36(1): 72-77, 85. JIANG Rong, YU Yi, TANG Yu-rong, et al. Effects of warming and biochar addition on soil enzyme activities in farmland[J]. Journal of Sichuan Agricultural University, 2018, 36(1): 72-77, 85. |
| [21] |
Fahad S, Hussain S, Saud S, et al. A combined application of biochar and phosphorus alleviates heat-induced adversities on physiological, agronomical and quality attributes of rice[J]. Plant Physiology and Biochemistry, 2016, 103: 191-198. |
| [22] |
OGuntunde P G, Abiodun B J, Ajayi A E, et al. Effects of charcoal production on soil physical properties in Ghana[J]. Journal of Plant Nutrition and Soil Science, 2008, 171(4): 591-596. |
| [23] |
Zhang Q Z, Wang Y D, Wu Y F, et al. Effects of biochar amendment on soil thermal conductivity, reflectance, and temperature[J]. Soil Science Society of America Journal, 2013, 77(5): 1478-1487. |
| [24] |
许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯, 1997, 24(4): 241-244. XU Da-quan. Some problems in stomatal limitation analysis of photosynthesis[J]. Plant Physiology Communications, 1997, 24(4): 241-244. |
| [25] |
高冠龙, 冯起, 张小由, 等. 植物叶片光合作用的气孔与非气孔限制研究综述[J]. 干旱区研究, 2018, 35(4): 929-937. GAO Guanlong, FENG Qi, ZHANG Xiao-you, et al. An overview of stomatal and non-stomatal limitations to photosynthesis of plants[J]. Arid Zone Research, 2018, 35(4): 929-937. |
| [26] |
张祎玮, 娄运生, 朱怀卫, 等. 夜间增温对水稻生长、生理特性及产量构成的影响[J]. 中国农业气象, 2017, 38(2): 88-95. ZHANG Yi-wei, LOU Yun-sheng, ZHU Huai-wei, et al. Impacts of night warming on rice growth, physiological properties and yield components[J]. Chinese Journal of Agrometeorology, 2017, 38(2): 88-95. |
| [27] |
杨泽粟, 张强, 郝小翠. 自然条件下半干旱雨养春小麦生育后期旗叶光合的气孔和非气孔限制[J]. 中国生态农业学报, 2015, 23(2): 174-182. YANG Ze-su, ZHANG Qiang, HAO Xiao-cui. Stomatal or non-stomatal limitation of photosynthesis of spring wheat flag leaf at late growth stages under natural conditions in semiarid rainfed regions[J]. Chinese Journal of Eco-Agriculture, 2015, 23(2): 174-182. |
| [28] |
吴冰洁, 刘玉军, 姜闯道, 等. 叶片生长进程中气孔发育对叶温调节的影响[J]. 植物生理学报, 2015, 51(1): 119-126. WU Bingjie, LIU Yu-jun, JIANG Chuang-dao, et al. Effects of stomatal development on leaf temperature during leaf expansion[J]. Plant Physiology Journal, 2015, 51(1): 119-126. |
| [29] |
张伟明, 孟军, 王嘉宇, 等. 生物炭对水稻根系形态与生理特性及产量的影响[J]. 作物学报, 2013, 39(8): 1445-1451. ZHANG Wei-ming, MENG Jun, WANG Jia-yu, et al. Effect of biochar on root morphological and physiological characteristics and yield in rice[J]. Acta Agronomica Sinica, 2013, 39(8): 1445-1451. |
| [30] |
Ma J F, Yamaji N. Silicon uptake and accumulation in higher plants[J]. Trends in Plant Science, 2006, 11(8): 392-397. |
| [31] |
韦还和, 孟天瑶, 李超, 等. 施硅量对甬优系列籼粳交超级稻产量及相关形态生理性状的影响[J]. 作物学报, 2016, 42(3): 437-445. WEI Huan-he, MENG Tian-yao, LI Chao, et al. Effects of silicon fertilizer rate on grain yield and related morphological and physiological characteristics in super rice of Yongyou japonica/indica hybrids series[J]. Acta Agronomica Sinica, 2016, 42(3): 437-445. |
| [32] |
徐呈祥, 刘兆普, 刘友良. 硅在植物中的生理功能[J]. 植物生理学通讯, 2004, 40(6): 753-757. XU Cheng-xiang, LIU Zhao-pu, LIU You-liang. The physiological function of silicon in plants[J]. Plant Physiology Communications, 2004, 40(6): 753-757. |
| [33] |
陈根云, 陈娟, 许大全. 关于净光合速率和胞间CO2浓度关系的思考[J]. 植物生理学通讯, 2010, 46(1): 64-66. CHEN Gen-yun, CHEN Juan, XU Da-quan. Thinking about the relationship between net photosynthetic rate and intercellular CO2 concentration[J]. Plant Physiology Communications, 2010, 46(1): 64-66. |
| [34] |
陈心想, 何绪生, 耿增超, 等. 生物炭对不同土壤化学性质、小麦和糜子产量的影响[J]. 生态学报, 2013, 33(20): 6534-6542. CHEN Xin-xiang, HE Xu-sheng, GENG Zeng-chao, et al. Effects of biochar on selected soil chemical properties and on wheat and millet yield[J]. Acta Ecologica Sinica, 2013, 33(20): 6534-6542. |
| [35] |
曹逼力, 徐坤, 石健, 等. 硅对番茄生长及光合作用与蒸腾作用的影响[J]. 植物营养与肥料学报, 2013, 19(2): 354-360. CAO Bili, XU Kun, SHI Jian, et al. Effects of silicon on growth, photosynthesis and transpiration of tomato[J]. Journal of Plant Nutrition and Fertilizers, 2013, 19(2): 354-360. |
| [36] |
李晓, 冯伟, 曾晓春. 叶绿素荧光分析技术及应用进展[J]. 西北植物学报, 2006, 26(10): 2186-2196. LI Xiao, FENG Wei, ZENG Xiao-chun. Advances in chlorophyll fluorescence analysis and its uses[J]. Acta Botanica Boreali-Occidentalia Sinica, 2006, 26(10): 2186-2196. |
| [37] |
Brestic M, Zivcak M, Kalaji H M, et al. Photosystem Ⅱ thermostability in situ: Environmentally induced acclimation and genotype-specific reactions in Triticum aestivum L.[J]. Plant Physiology & Biochemistry, 2012, 57(8): 93-105. |
| [38] |
Rai M K, Shend S, Strasser R J. JIP test for fast fluorescence transients as a rapid and sensitive technique in assessing the effectiveness of arbuscular mycorrhizal fungi in Zea mays: Analysis of chlorophyll a fluorescence[J]. Plant Biosystems, 2008, 142(2): 191-198. |
| [39] |
冯芳, 范佩佩, 刘超, 等. 水稻叶绿素荧光特性对CO2浓度升高的代际响应研究[J]. 生态环境学报, 2019, 28(3): 463-471. FENG Fang, FAN Pei-pei, LIU Chao, et al. Intergenerational response of chlorophyll fluorescence characteristics of rice to elevated CO2 concentration[J]. Ecology and Environmental Sciences, 2019, 28(3): 463-471. |
| [40] |
张国良, 戴其根, 王建武, 等. 施硅量对粳稻品种武育粳3号产量和品质的影响[J]. 中国水稻科学, 2007(3): 299-303. ZHANG Guo-liang, DAI Qi-gen, WANG Jian-wu, et al. Effects of silicon fertilizer rate on yield and quality of japonica rice Wuyujing 3[J]. Chinese Journal of Rice Science, 2007(3): 299-303. |