 2021, Vol. 40
2021, Vol. 40



池塘养殖是我国重要的水产养殖方式。近30 a来,我国池塘养殖规模和产量呈不断增加趋势。2018年,全国淡水池塘养殖面积达2.67×106 hm2,养殖产量达2.21×1010 kg,分别占淡水养殖总面积和总产量的51.8% 和74.7%[1]。为了提高产量,我国池塘养殖普遍采用高密度、高投饵率的精养模式[2],过量的饲料投入和鱼类代谢产物的积累导致养殖塘体氮等养分污染严重。以往研究显示,鱼、虾等水产品生长所吸收的氮仅占饵料投入的20.0%~27.2%,超过70%的氮残留在养殖塘底泥和水体中。其中底泥中的氮富集尤为严重,占总投入量的50.4%~69.6%[3-5]。养殖池塘冗余的氮不仅会影响鱼虾的正常生长,尾水的排放还会对外源水体产生富营养化污染[6]。近年来,养殖池塘N2O排放问题也日益引起关注[7-9]。全球数据的估算显示,到2030年全球水产养殖N2O年排放量将达到3.83×108 kg,占人类总排放的5.7%,是主要的人为排放源之一[10]。如何消减养殖塘体的N2O排放,是当前池塘养殖可持续发展面临的重要问题之一。
作物(水稻、蔬菜等)-鱼共作是我国典型的生态农作模式。以往研究显示[11-13],稻-鱼共作或鱼-菜共作有利于提高农作系统氮磷等养分利用效率、减少养分流失。近年来,有研究开始关注稻-鱼共作对养殖水体N2O排放的影响。Li等[14]研究显示,稻鱼共作养殖池塘比单养鱼养殖池塘N2O排放量和排放系数分别降低85.6% 和83.3%。Datta等[15]对稻田养鱼模式的研究显示,与水稻单作相比,稻鱼共作模式可以减少19.4% 的N2O排放。对于鱼-菜共生系统N2O排放的研究则相对较少,邹艺娜等[16]报道了在鱼菜共生系统中,以N2O形式释放的氮素约占氮素总投入的1.54%,与普通水产养殖相比,其N2O转化率没有明显增加。不同作物由于生长特征和生物量的差异,在与水产动物共作中,对系统中氮素转化的调控可能存在较大差异。综合比较不同作物对养殖水体冗余氮素的富集效应以及对N2O排放的消减效应,可以为共生系统中作物的选择提供理论依据和参考。
目前对作物-鱼共作系统N2O产生和排放的作用机理研究还非常有限。反硝化过程是养殖系统中N2O产生的主要途径之一[17]。反硝化作用包括4个步骤:NO3--NO2--NO-N2O-N2,此过程中的酶分别为硝酸还原酶(Nar)、亚硝酸还原酶(Nir)、NO还原酶(Nor)和N2O还原酶(Nos)[10]。亚硝酸盐转化为氧化氮过程是反硝化作用的标志性反应,控制关键的限速步骤[18]。nirK和nirS基因是两种不同类型的亚硝酸盐还原酶基因,nosZ基因是唯一编码N2O还原酶的基因,因此nirK、nirS、nosZ等基因是常用的检测反硝化微生物群落的功能基因[19]。因此,研究作物-鱼共作系统对nirK、nirS、nosZ等关键功能基因的影响,可以为揭示其对N2O产生和排放的作用机理提供重要参考。
本研究通过盆栽模拟试验,研究4种作物[粳稻(Oryza sativa L. subsp. japonica Kato)、籼稻(Oryza sativa L. subsp. sativa)、小白菜(Brassica rapa L. ssp. pekinensis)和空心菜(Ipomoea aquatic Forsk)] 与黄颡鱼(Pelteobagrus fulvidraco)共作对养殖水体N2O排放通量、功能基因、养殖水体和底泥氮浓度,以及对氮素利用效率差异的影响,以期能为共生系统中作物品种的选择提供理论支持。
1 材料与方法 1.1 试验设计试验于2019年在浙江省杭州市中国水稻研究所实验基地(119.95°E,30.05°N)进行。试验设置了5个处理:粳稻(安粳优1号)+黄颡鱼(JR-F)、籼稻(农两优渔1号)+黄颡鱼(IR-F)、小白菜(杂交青梗)+黄颡鱼(LC-F)、空心菜(柳叶空心菜)+黄颡鱼(WS-F)和单养黄颡鱼对照(CK)。每个处理3个重复。采用盆栽模拟试验,试验选用长宽高分别为1.75、1.25、1 m的塑料大桶,以保证水产动物有充足的活动空间。在试验基地附近选择了一个连续养殖10 a的精养池塘,挖取池塘底泥,混匀后,铺在盆栽塑料大桶底部,底泥铺设高度为20 cm,以模拟池塘环境。
4种作物均于2019年6月21日移栽,粳稻、籼稻、小白菜和空心菜的初始生物量(干质量)分别为10.66、11.59、21.11 g·m-2和6.69 g·m-2。水稻直接种植于底泥中,每个桶栽种4丛水稻,每丛2株,株行距为50 cm×50 cm。空心菜和小白菜采用浮床种植,每个桶设置4个浮床,浮床尺寸为0.4 m×0.4 m。每个浮床种植9株蔬菜,株行距8 cm×8 cm。作物移栽40 d后,开始放养黄颡鱼苗,CK的鱼苗放养时间与共作系统相同。非共作期为作物单作期间,时间为6月21日—7月31日,共作期为水稻/蔬菜-鱼共作时期,时间为8月1日—10月23日。放养规格为4~5 g·尾-1,放养量为80尾·桶-1。放鱼后使用160 W增氧泵每日上午9:00—11:00、下午14:00—18:00、晚上23:00—次日早上6:00桶内曝气增氧,取样的上午暂停增氧。随着水稻不断生长,每周添加去氯后的自来水逐渐提高水位。桶内水位最高为85 cm。所有处理的水位保持一致。试验期间未使用化学肥料和农药。黄颡鱼驯养3 d后投喂配合饲料,日投饲两次,分别为上午7:00和下午6:00;日投饲量为黄颡鱼体质量的4%,上午占40%,下午占60%。整个试验周期氮投入量为150.83 g。根据黄颡鱼体质量、食用量和反应及时增添饲料。水稻于2019年10月23日收获;蔬菜每月收获1次,最后一次收获时间为10月23日,共收获4次;黄颡鱼在水稻收获后第2 d捕获。
1.2 采样与分析 1.2.1 N2O净排放通量N2O净排放通量采用静态箱-气相色谱法测定[20]。静态箱规格为0.5 m×0.5 m×0.5 m。采样箱内配有风扇以确保气体完全混合,外覆保温和反光材料,减少采样期间外界温度和太阳光的干扰。每个试验桶内安装可调节高度的支架,取样时将静态箱放置在支架上进行取气。在整个试验期,每周一次,在上午8:00—10:00用铝箔气袋以10 min的间隔采集气体样品。气体样品带回实验室后,采用气相色谱仪(GC 2010,岛津)分析气体样品中N2O浓度。N2O排放通量采用公式(1)计算[14]:

|
(1) |
式中:F为N2O排放通量,μg·m-2·h-1;ρ为标准状态下N2O密度,1.964 kg·m-3;V为静态箱的体积,m3;A为静态箱的基底面积,m2;P为箱内大气压,P0为标准大气压,kPa;dC/dt为采样箱内N2O浓度随时间变化的斜率,10-9 min-1;T为采样过程中静态箱内平均温度,℃。
1.2.2 土壤孔隙水和上覆水N2O浓度在水稻移栽后,将直径0.5 cm、长度5 cm的陶瓷土壤溶液采样器垂直插入土壤中,用20 mL注射器采集上覆水(水面以下20 cm)和土壤孔隙水10 mL,注入20 mL真空玻璃瓶中。每月采集一次。样品采集后立刻运回实验室,用顶空-平衡法测定上覆水和土壤孔隙水中N2O浓度。具体过程为:先将样品平衡温度至室温,使用注射器于样品玻璃瓶中注入N2以中和压力,剧烈摇动样品并使其平衡20 min。采用注射器将玻璃瓶中气体吸入,将其注入气相色谱仪中,测定N2O的浓度。土壤孔隙水和上覆水中N2O浓度采用公式(2)计算[21-23]:
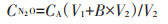
|
(2) |
式中:CN2O为上覆水/土壤孔隙水中N2O浓度,μL·L-1;CA为真空玻璃瓶中顶空气体N2O浓度,μL·L-1;V1和V2分别为真空玻璃瓶中气体和液体的体积,L;B为Bunsen溶解系数(26 ℃)。
1.2.3 水体和底泥养分等常规指标和功能基因测定在气体采样的同一天采用取水器采集每个处理的养殖水体样品。在每个桶里采集两个20 cm深处水样500 mL混合均匀后作为一个混合样品,水样带回实验室后4 ℃保存,24 h内测定。水样经0.45 μm玻璃纤维滤膜过滤后,采用连续流动分析仪(Skalar San++,荷兰)测量水中铵态氮(NH4+-N)、硝态氮(NO3--N)、亚硝态氮(NO2--N)的浓度。总氮(TN)采用过硫酸钾氧化法测定;溶氧(DO)和pH通过梅特勒-托利多溶氧仪S7和手持式pH仪S2原位测量。
采用底泥取样器采集每个处理的底泥样品,每月一次。在每个桶里采集3个表层(0~5 cm)底泥样品,混匀后分为3份,一份自然风干,另一份-20 ℃保存,第三份-80 ℃保存测微生物功能基因。风干样采用凯氏定氮法测TN;鲜样冷冻干燥后采用靛酚蓝比色法测NH4+-N和采用紫外分光光度法测NO3--N;鲜样采用水提取-过硫酸钾氧化法测定可溶性有机氮(DON)[24]。
反硝化过程重要的酶基因[土壤亚硝酸盐还原酶(nirK、nirS)和N2O还原酶(nosZ)]丰度采用实时荧光定量PCR分析。即用土壤DNA快速提取试剂盒(MP Biomedicals,美国)提取土壤DNA,用0.8% 的琼脂糖凝胶电泳检测所提DNA片段大小,并用超微量紫外分光光度计测定DNA浓度及质量[25]。设计nirK、nirS、nosZ基因引物,进行实时荧光定量PCR分析。
2 结果与分析 2.1 N2O排放全试验期不同作物-鱼共作和单养鱼处理N2O的排放通量动态变化如图 1所示。各处理N2O排放通量呈波动变化趋势。单养鱼处理(CK)在7月8日和10月14日出现较高的N2O排放峰,其他处理N2O排放峰相对较低(图 1)。进一步分析了单养鱼处理出现N2O排放峰时水体NO3--N、NO2--N和DO含量特征(图 2),在7月8日出现第一个N2O排放峰时,养殖水体NO3--N和NO2--N含量均出现峰值;而10月14日出现第二个N2O排放峰时,除了水体NO3--N含量较高外,DO含量也较低。这说明N2O排放峰的出现可能与水体NO3--N和NO2--N含量较高以及DO含量较低有关。

|
图 1 不同处理全试验期N2O排放通量的动态变化 Figure 1 Dynamic change of N2O emission flux of different treatments during full experimental stage |

|
图 2 单养鱼处理全试验期养殖水体NO3--N、NO2--N、DO浓度和N2O排放通量的变化 Figure 2 Variation of NO3--N, NO2--N and DO concentrations in the aquaculture water and N2O emission flux in fish monoculture system during full experimental stage |
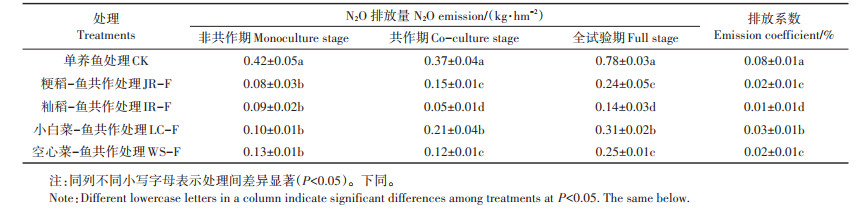
在非共作期(6月21日—7月31日)、共作期(8月1日—10月21日)以及全试验期,单养鱼处理N2O排放量和排放系数均显著高于4种作物-鱼共作处理(表 1)。在非共作期,4种作物-鱼共作处理之间N2O排放量没有显著差异。但是在共作期,籼稻-鱼共作处理N2O排放量要显著低于蔬菜-鱼共作处理。从全试验期来看,在4种共作处理中,籼稻-鱼共作处理对N2O的消减效应最佳。与单养鱼处理相比,其N2O排放总量减少了82.1%。其次是粳稻-鱼和空心菜-鱼处理,其N2O排放总量分别减少了69.2%和67.9%;小白菜-鱼共作处理对N2O的消减效应相对较低,其N2O排放总量仅减60.3%。
|
|
表 1 不同处理非共作期、共作期和全试验期的N2O排放量和排放系数 Table 1 N2O emission amount and emission coefficient of different treatments during monoculture stage, co-culture stage and full experimental stage |
上覆水和土壤孔隙水中N2O浓度均值如图 3所示。不同处理上覆水N2O浓度在0.75~1.36 μL·L-1。作物-鱼共作处理上覆水N2O浓度均显著低于单养鱼,JR-F、IR-F、LC-F和WS-F处理分别比CK处理降低30.0%、43.3%、24.4% 和44.4%。不同处理土壤孔隙水中N2O浓度在0.54~1.92 μL·L-1。单养鱼处理上覆水N2O浓度和土壤孔隙水浓度无显著差异;粳稻和籼稻-鱼共作处理土壤孔隙水N2O浓度均低于上覆水(图 3),分别减少了37.1% 和24.2%;而小白菜和空心菜-鱼共作处理土壤孔隙水N2O浓度均显著高于上覆水中N2O浓度,分别增加了87.1%和146.6%。

|
不同小写字母表示处理间差异显著(P < 0.05)。下同 Different lowercase letters indicate significant differences among treatments at P < 0.05. The same below 图 3 不同处理全试验期上覆水和土壤孔隙水N2O浓度均值 Figure 3 Average N2O concentrations in the overlying water and soil pore water of different treatments during full experimental stage |
与上覆水中N2O规律不同,两种蔬菜-鱼共作处理土壤孔隙水N2O浓度与单养鱼处理没有显著差异。两种水稻-鱼共作处理土壤孔隙水N2O浓度则分别比单养鱼处理降低了61.2%和62.1%。这与水稻和蔬菜的种植方式有关。蔬菜种植在浮床上,仅能减少养殖水体中N2O浓度,而不能消减土壤孔隙水中N2O浓度。种植水稻可以同时减少养殖水体和土壤孔隙水中N2O浓度。
2.3 养殖水体和底泥氮浓度非共作期、共作期和全试验期各处理养殖水体氮浓度如图 4所示。在非共作期,与单养鱼处理相比,两种水稻-鱼共作处理显著降低养殖水体TN、NH4+-N、NO3--N和NO2--N浓度;两种蔬菜-鱼共作处理显著降低养殖水体NO3--N浓度,TN、NH4+-N、NO2--N浓度与单养鱼处理没有显著差异。在共作期,两种水稻-鱼共作处理养殖水体TN、NH4+-N、NO3--N、NO2--N浓度均与单养鱼处理没有显著差异,两种蔬菜-鱼共作处理显著降低养殖水体TN、NO3--N浓度。从全试验期来看,两种水稻-鱼共作处理显著降低养殖水体TN、NH4+-N、NO3--N浓度,NO2--N浓度与单养鱼没有显著差异;两种蔬菜-鱼共作养殖水体显著降低NO3--N浓度,TN、NH4+-N、NO2--N浓度与单养鱼处理没有显著差异。

|
图 4 不同处理非共作期、共作期和全试验期养殖水体TN、NH4+-N、NO3--N、NO2--N浓度 Figure 4 Concentrations of TN, NH4+-N, NO3--N and NO2--N in the water of different treatments during monoculture stage, co-culture stage |
各处理底泥氮含量如图 5所示。在非共作期,4种作物-鱼共作处理底泥TN、NH4+-N、NO3--N、DON含量与单养鱼处理没有显著差异,但是两种水稻-鱼共作处理NH4+-N、DON含量显著低于两种蔬菜-鱼共作处理。在共作期,两种水稻-鱼共作处理显著降低了底泥TN、NH4+-N、DON含量,NO3--N含量与单养鱼处理没有显著差异;两种蔬菜-鱼共作处理底泥TN、NH4+-N、NO3--N、DON含量均与单养鱼没有显著差异。在全试验期,两种水稻-鱼共作处理显著降低了底泥NH4+-N和DON含量;两种蔬菜-鱼共作处理底泥TN、NH4+-N、NO3--N、DON含量与单养鱼没有显著差异。这些结果表明,两种水稻-鱼共作处理可以减少底泥NH4+-N和DON浓度,共作期的消减效应较强;两种蔬菜-鱼共作处理不能减少底泥氮浓度。

|
图 5 不同处理非共作期、共作期和全试验期底泥TN、NH4+-N、NO3--N和DON浓度 Figure 5 Concentrations of TN, NH4+-N, NO3--N and DON in soil of different treatments during monoculture stage, co-culture stage and full experimental stage |
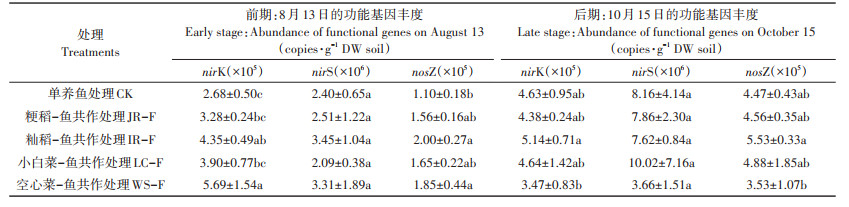
分别测定了不同处理共作处理前期(8月13日)和后期(10月15日)底泥中反硝化细菌基因丰度,如表 2所示。前期不同作物-鱼共作处理土壤亚硝酸盐还原酶(nirK、nirS)和N2O还原酶(nosZ)基因丰度均小于后期。前期单养鱼处理土壤nirK基因丰度最低,显著低于籼稻-鱼和空心菜-鱼共作处理;4种作物-鱼共作处理与单养鱼处理底泥nirS基因丰度无显著差异;底泥nosZ基因丰度与nirK基因丰度规律一致,同样表现为单养鱼处理土壤nosZ基因丰度最小,且显著低于籼稻-鱼和空心菜-鱼共作处理。后期单养鱼处理土壤亚硝酸盐还原酶(nirK、nirS)和N2O还原酶(nosZ)基因丰度与4种作物-鱼共作处理都没有显著差异,但是籼稻-鱼共作处理土壤nirK和nosZ基因丰度均显著高于空心菜-鱼共作处理。这些结果表明,4种作物-鱼共作和单养鱼处理的功能基因差异主要体现在nirK和nosZ基因丰度上,籼稻-鱼共作处理前期和后期nosZ基因丰度均最高,促进反硝化中间产物N2O还原为N2,与籼稻-鱼共作处理N2O排放总量最少规律一致。
|
|
表 2 不同处理共作前期和后期底泥nirK、nirS、nosZ基因丰度 Table 2 Gene abundance of nirK, nirS and nosZ in soil of different treatments at early and late co-culture stages |
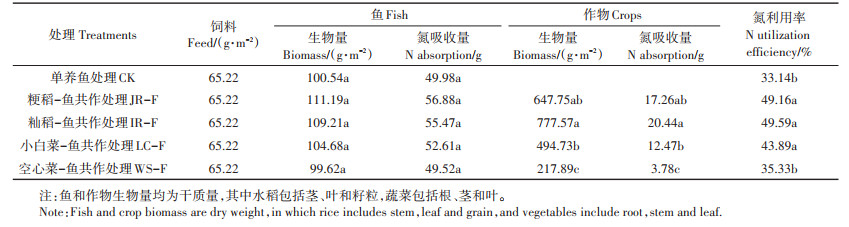
2019年盆栽试验的氮投入均为饲料投入。试验结束后4种作物-鱼共作处理收获的黄颡鱼生物量和氮吸收量与单养鱼处理没有显著差异(表 3);作物收获后,两种水稻的生物量和氮吸收量均高于两种蔬菜,其中空心菜的生物量与氮吸收量最低。4种作物-鱼共作处理氮利用率均高于单养鱼处理,其中籼稻-鱼、粳稻-鱼和小白菜-鱼共作处理氮利用率均显著增加。
|
|
表 3 不同处理鱼和作物的生物量、氮吸收量以及氮利用率 Table 3 Biomass, N absorption and N utilization efficiency of fish and crops under different treatments |
淡水养殖系统N2O排放可能受到水生植物、水体无机氮浓度、DO浓度、温度、pH等因素的影响[26-27]。以往研究发现,N2O排放峰易出现在高无机氮浓度、低DO浓度条件下[7, 28]。本研究发现单养鱼处理在7月8日和10月14日出现较高的N2O排放峰,其他处理N2O排放峰相对较低(图 1)。通过将单养鱼处理N2O通量与养殖水体NO3--N、NO2--N和DO含量对比发现,7月8日养殖水体NO3--N和NO2--N含量均出现峰值(图 2),这可能主要因为CK处理没有种植水稻,前期水体和底泥中存在大量无机氮,经硝化和反硝化作用产生N2O。因此,充沛的无机氮底物可能是7月8日测量出现N2O排放峰的主要原因。在后期,10月14日出现N2O排放峰,可能有两方面的原因,一是充沛的底物,10月14日前后水体NO3--N含量也较高;二是10月14日水体DO较低,这可能是连续阴雨天,再加上后期CK处理水质的恶化,藻类呼吸耗氧以及有机物降解耗氧导致水体DO下降。
如何消减池塘养殖的N2O排放问题日益引起关注。本研究选取了两种水稻(粳稻和籼稻)和两种蔬菜(小白菜和空心菜)作为研究对象,比较不同类型作物-鱼共作对养殖系统N2O排放的影响。研究结果显示,4种作物-鱼共作处理均显著降低了N2O排放总量和排放系数。这与Li等[14]、袁伟玲等[29]对稻-鱼共作系统的研究结果相近。但是Sumeth等[30]的研究结果表明罗非鱼-小白菜、生菜、韭菜和西红柿等水培浮床系统的N2O排放通量高于单养鱼处理。这可能是因为该研究的系统结构与本研究不同,其研究的共作系统是鱼池、浮床、过滤池和曝气池联合,排放总量由多个系统叠加,而本研究的作物和鱼培养在同一生态系统中。也有研究表明,养殖池塘种植伊乐藻、轮叶黑藻等水生植物对N2O排放无显著影响[31]。因此,作物-鱼共作对养殖系统N2O排放的影响与研究品种和系统结构有关。
作物-鱼共作对养殖系统N2O的消减可能存在多种途径。一方面,水稻和蔬菜的生长可以直接吸收底泥或水体中的无机氮。与单养鱼相比,水稻-鱼共作可以显著降低底泥NH4+-N和DON浓度,显著降低水体TN、NH4+-N、NO3--N浓度;蔬菜-鱼共作可以显著降低水体NO3--N浓度(图 4和图 5)。因此,水稻和蔬菜的生长吸收减少了N2O产生的底质氮[14],从而减少N2O的排放。另一方面,在养殖系统中,反硝化作用是底泥产生N2O的关键途径[32],反硝化微生物功能基因(nirK、nirS和nosZ)丰度决定了N2O的排放潜力[33]。作物与鱼共作,作物的通气组织可以通过蒸腾作用和气体交换将氧气从大气中转移到根部区域[34],从而提高根系区域氧气的浓度[35],同时根系呼吸消耗氧气,在根系部分形成根际好氧、兼氧和厌氧区域,为不同类型的微生物提供良好生存环境[36]。好氧微生物对有机质的分解促进反硝化微生物的生长,厌氧区域也有利于反硝化微生物的生长和繁殖[37],功能基因的测定结果也显示,作物-鱼共作有利于提高底泥中nirK和nosZ功能基因丰度(表 2)。因此,作物-鱼共作有利于反硝化微生物的生长繁殖,加速反硝化速率,促进N2O进一步还原为N2,减少N2O的排放。
本研究发现,两种水稻-鱼共作处理的减排效果显著优于两种蔬菜-鱼共作处理。其原因可能主要是作物种植方式的差异。水稻与鱼共作,是选用高秆品种直接种植在底泥中,水稻的生长可以同时吸收水体(有水生根)和底泥中无机氮(图 4和图 5),抑制水体和底泥中N2O的产生(图 3);而蔬菜与鱼共作是种植在浮床上,仅能降低养殖水体氮元素(图 4),对底泥中无机氮没有显著影响(图 5)。因此,其对N2O的消减作用要弱于水稻。在两种水稻-鱼共作处理中,籼稻-鱼共作处理的减排效应显著优于粳稻-鱼共作处理。这可能主要有两方面的原因:一方面,籼稻的生物量高于粳稻(表 3),籼稻吸收和利用的氮素较多。籼稻-鱼共作处理非共作期养殖水体TN、NH4+-N浓度、底泥DON浓度和共作期养殖水体NH4+-N、NO2--N浓度、底泥NH4+-N、DON浓度均低于粳稻-鱼共作处理(图 4和图 5)。作物吸收和利用的氮素较多,而供给微生物的氮素则相应减少。因此籼稻对于产N2O底质氮的消减作用要强于粳稻。另一方面,籼稻-鱼共作处理土壤nosZ基因丰度在共作初期和后期均高于粳稻-鱼共作处理(表 2),可能提高了N2O还原为N2的能力,导致反硝化作用产生的N2O总量减少。对于两种蔬菜-鱼共作处理,空心菜-鱼共作处理N2O排放量显著低于小白菜-鱼处理(表 1)。虽然小白菜的生物量和氮吸收量均高于空心菜(表 3),但在全试验期小白菜-鱼共作和空心菜-鱼共作处理水体总氮和无机氮含量均没有显著差异(图 4和图 5)。因此,两种蔬菜-鱼共作处理对N2O排放效应的差异可能主要是由于两种蔬菜对反硝化过程的功能基因以及N2O排放过程的影响存在差异。其作用机理还需要进一步深入研究。
4 结论与单养鱼处理相比,作物-鱼共作处理均显著降低淡水养殖系统N2O的排放。在共作处理N2O产生过程中,作物对底泥中的nirK和nosZ功能基因丰度均有显著调控作用,籼稻-鱼共作处理前期和后期nosZ基因丰度均最高,显著减少了上覆水和土壤孔隙水中N2O浓度。同时作物-鱼共作显著降低养殖水体和底泥氮浓度,减少水产养殖氮污染。作物-鱼共作处理获得了水产品和作物产品,提高了氮素利用率。这些结果表明,作物-鱼共作具有减缓N2O排放的潜力,是减轻气态氮损失的有效途径。
| [1] |
农业农村部渔业渔政管理局. 2019年中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2019: 21-49. Fisheries and Fisheries Administration of the Ministry of Agriculture and Rural Affairs. 2019 China fishery statistical yearbook[M]. Beijing: China Agriculture Press, 2019: 21-49. |
| [2] |
臧维玲, 姚庆祯, 戴习林, 等. 上海地区水产养殖和长江口与杭州湾水域环境的关系[J]. 上海水产大学学报, 2003(3): 219-226. ZANG Wei-ling, YAO Qing-zhen, DAI Xi-lin, et al. The relationship between the aquaculture in Shanghai area and water environment in the mouth of Yangtse River and Hangzhou Bay[J]. Journal of Shanghai Fisheries University, 2003(3): 219-226. |
| [3] |
Green B W, Boyd C E. Chemical budgets for organically fertilized fish ponds in the dry tropics[J]. Journal of the World Aquaculture Society, 1995, 26(3): 284-296. DOI:10.1111/j.1749-7345.1995.tb00257.x |
| [4] |
刘兴国. 池塘养殖污染与生态工程化调控技术研究[D]. 南京: 南京农业大学, 2011: 17-24. LIU Xing-guo. Study on the pond aquaculture pollution and ecological engineering regulation techniques[D]. Nanjing: Nanjing Agricultural University, 2011: 17-24. |
| [5] |
李卓佳, 虞为, 朱长波, 等. 对虾单养和对虾-罗非鱼混养试验围隔氮磷收支的研究[J]. 安全与环境学报, 2012, 12(4): 50-55. LI Zhuo-jia, YU Wei, ZHU Chang-bo, et al. Study on the nitrogen and phosphorus budgets of experimental enclosures with shrimp monoculture and shrimp-tilapia polyculture[J]. Journal of Safety and Environment, 2012, 12(4): 50-55. DOI:10.3969/j.issn.1009-6094.2012.04.012 |
| [6] |
Li X P, Li J R, Wang Y B, et al. Aquaculture industry in China: Current state, challenges, and outlook[J]. Reviews in Fisheries Science, 2011, 19(3): 187-200. DOI:10.1080/10641262.2011.573597 |
| [7] |
Hu Z, Lee W S, Chandran K, et al. Nitrogen transformations in intensive aquaculture system and its implication to climate change through nitrous oxide emission[J]. Bioresource Technology, 2013, 130: 314-320. DOI:10.1016/j.biortech.2012.12.033 |
| [8] |
Yogev U, Atari A, Gross A. Nitrous oxide emissions from near-zero water exchange brackish recirculating aquaculture systems[J]. Science of the Total Environment, 2018, 628/629: 603-610. DOI:10.1016/j.scitotenv.2018.02.089 |
| [9] |
Hu Z, Lee J W, Chandran K, et al. Influence of carbohydrate addition on nitrogen transformations and greenhouse gas emissions of intensive aquaculture system[J]. Science of the Total Environment, 2014, 470/471: 193-200. DOI:10.1016/j.scitotenv.2013.09.050 |
| [10] |
Hu Z, Lee J W, Chandran K, et al. Nitrous oxide(N2O)emission from aquaculture: A review[J]. Environmental Science & Technology, 2012, 46(12): 6470-6480. |
| [11] |
李凤博, 冯金飞, 周锡跃, 等. 鱼塘种稻对养殖水体营养物质的去除作用研究[J]. 中国水稻科学, 2015, 29(2): 174-180. LI Fengbo, FENG Jin-fei, ZHOU Xi-yue, et al. Nutrients removal from fish pond by rice planting[J]. Chinese Journal of Rice Science, 2015, 29(2): 174-180. DOI:10.3969/j.issn.1001-7216.2015.02.009 |
| [12] |
李成芳, 曹凑贵, 汪金平, 等. 稻鸭稻鱼共作生态系统N素平衡的研究[J]. 农业环境科学学报, 2008, 27(4): 1326-1334. LI Chengfang, CAO Cou-gui, WANG Jin-ping, et al. Studies on nitrogen cycling in integrated rice-duck, rice-fish ecosystems[J]. Journal of Agro-Environment Science, 2008, 27(4): 1326-1334. DOI:10.3321/j.issn:1672-2043.2008.04.009 |
| [13] |
Hu Z, Lee J W, Chandran K, et al. Effect of plant species on nitrogen recovery in aquaponics[J]. Bioresource Technology, 2015, 188: 92-98. DOI:10.1016/j.biortech.2015.01.013 |
| [14] |
Li F B, Feng J F, Zhou X Y, et al. Impact of rice-fish/shrimp co-culture on the N2O emission and NH3 volatilization in intensive aquaculture ponds[J]. Science of the Total Environment, 2019, 655: 284-291. DOI:10.1016/j.scitotenv.2018.10.440 |
| [15] |
Datta A, Nayak D R, Sinhababu D P, et al. Methane and nitrous oxide emissions from an integrated rainfed rice–fish farming system of Eastern India[J]. Agriculture, Ecosystems & Environment, 2009, 129(1/2/3): 228-237. |
| [16] |
邹艺娜, 胡振, 张建, 等. 鱼菜共生系统氮素迁移转化的研究与优化[J]. 环境工程学报, 2015, 9(9): 4211-4216. ZOU Yi-na, HU Zhen, ZHANG Jian, et al. Investigation and optimization of nitrogen transformations in aquaponics[J]. Chinese Journal of Environment Engineering, 2015, 9(9): 4211-4216. |
| [17] |
Wrage N, Lauf J, A Prado A, et al. Distinguishing sources of N2O in European grasslands by stable isotope analysis[J]. Rapid Commun Mass Spectrom, 2004, 18(11): 1201-1207. DOI:10.1002/rcm.1461 |
| [18] |
Braker G, Zhou J Z, Wu L Y, et al. Nitrite reductase genes(nirK and nirS)as functional markers to investigate diversity of denitrifying bacteria in pacific northwest marine sediment communities[J]. Applied and Environmental Microbiology, 2000, 66(5): 2096-2104. DOI:10.1128/AEM.66.5.2096-2104.2000 |
| [19] |
Levy-Booth D J, Prescott C E, Grayston S J. Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems[J]. Soil Biology & Biochemistry, 2014, 75: 11-25. |
| [20] |
Liu S W, Hu Z Q, Wu S, et al. Methane and nitrous oxide emissions reduced following conversion of rice paddies to inland crab-fish aquaculture in southeast China[J]. Environmental Science & Technology, 2016, 50(2): 633-642. |
| [21] |
Cheng W G, Yagi K, Sakai H, et al. Changes in concentration and δ13C value of dissolved CH4, CO2 and organic carbon in rice paddies under ambient and elevated concentrations of atmospheric CO2[J]. Organic Geochemistry, 2005, 36(5): 813-823. DOI:10.1016/j.orggeochem.2005.01.009 |
| [22] |
Terry R E, Tate R L, Duxbury J M. The effect of flooding on nitrous oxide emissions from an organic soil[J]. Soil Science, 1981, 132(3): 228-232. DOI:10.1097/00010694-198109000-00006 |
| [23] |
Weiss R F, Price B A. Nitrous oxide solubility in water and seawater[J]. Marine Chemistry, 1980, 8(4): 347-359. DOI:10.1016/0304-4203(80)90024-9 |
| [24] |
鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2020: 125-135. LU Ru-kun. Analytical methods for agrochemistry of soils[M]. Beijing: China Agricultural Science and Technology Press, 2020: 125-135. |
| [25] |
许欣, 陈晨, 熊正琴. 生物炭与氮肥对稻田甲烷产生与氧化菌数量和潜在活性的影响[J]. 土壤学报, 2016, 53(6): 1517-1527. XU Xin, CHEN Chen, XIONG Zheng-qin. Effects of biochar and nitrogen fertilizer amendment on abundance and potential activity of methanotrophs and methanogens in paddy field[J]. Acta Pedologica Sinica, 2016, 53(6): 1517-1527. |
| [26] |
丁维新, 袁俊吉, 刘德燕, 等. 淡水养殖系统温室气体CH4和N2O排放量研究进展[J]. 农业环境科学学报, 2020, 39(4): 749-761. DING Wei-xin, YUAN Jun-jie, LIU De-yan, et al. CH4 and N2O emissions from freshwater aquaculture[J]. Journal of Agro-Environment Science, 2020, 39(4): 749-761. |
| [27] |
安桐彤, 许潇方, 高强, 等. 我国养殖水体N2O排放特征及其影响因素研究进展[J]. 生态学杂志, 2021, 40(1): 266-277. AN Tongtong, XU Xiao-fang, GAO Qiang, et al. Spatiotemporal distribution and the influencing factors of nitrous oxide emissions from aquaculture water in China: A review[J]. Chinese Journal of Ecology, 2021, 40(1): 266-277. |
| [28] |
Ma Y C, Sun L Y, Liu C Y, et al. A comparison of methane and nitrous oxide emissions from inland mixed-fish and crab aquaculture ponds[J]. Science of the Total Environment, 2018, 637/638: 517-523. DOI:10.1016/j.scitotenv.2018.05.040 |
| [29] |
袁伟玲, 曹凑贵, 李成芳, 等. 稻鸭、稻鱼共作生态系统CH4和N2O温室效应及经济效益评估[J]. 中国农业科学, 2009, 42(6): 2052-2060. YUAN Wei-ling, CAO Cou-gui, LI Cheng-fang, et al. Methane and nitrous oxide emissions from rice-fish and rice duck complex ecosystems and the evaluation of their economic significance[J]. Scientia Agricultura Sinica, 2009, 42(6): 2052-2060. DOI:10.3864/j.issn.0578-1752.2009.06.022 |
| [30] |
Sumeth W, Brain N P, Samir K K. Nitrogen recovery and nitrous oxide (N2O)emissions from aquaponic systems: Influence of plant species and dissolved oxygen[J]. International Biodeterioration & Biodegradation, 2018, 134: 117-126. |
| [31] |
林海, 周刚, 李旭光, 等. 夏季池塘养殖中华绒螯蟹生态系统温室气体排放及综合增温潜势[J]. 水产学报, 2013, 37(3): 417-424. LIN Hai, ZHOU Gang, LI Xu-guang, et al. Greenhouse gases emissions from pond culture ecosystem of Chinese mitten crab and their comprehensive global warming potentials in summer[J]. Journal of Fisheries of China, 2013, 37(3): 417-424. |
| [32] |
Luo J, Tillman R W, Ball P R. Factors regulating denitrification in a soil under pasture[J]. Soil Biology & Biochemistry, 1999, 31(6): 913-927. |
| [33] |
Zhao S Y, Zhou J M, Yuan D D, et al. NirS-type N2O-producers and nosZ Ⅱ-type N2O-reducers determine the N2O emission potential in farmland rhizosphere soils[J]. Journal of Soils and Sediments, 2019, 20(1): 461-471. DOI:10.1007/s11368-019-02395-3 |
| [34] |
Colmer T D, Cox M C, Voesenek L A C. Root aeration in rice(Oryza sativa): Evaluation of oxygen, carbon dioxide, and ethylene as possible regulators of root acclimatizations[J]. New Phytologist, 2006, 170(4): 767-777. DOI:10.1111/j.1469-8137.2006.01725.x |
| [35] |
邹国元, 张福锁. 根际反硝化作用与N2O释放[J]. 中国农业大学学报, 2002, 7(1): 77-82. ZOU Guo-yuan, ZHANG Fu-suo. Denitrification in rhizosphere and N2O emission[J]. Journal of China Agriculturral University, 2002, 7(1): 77-82. |
| [36] |
Fennessy M S, Cronk J K, Mitsch W J. Macrophyte productivity and community development in created freshwater wetlands under experimental hydrological conditions[J]. Ecological Engineering, 1994, 3(4): 469-484. DOI:10.1016/0925-8574(94)00013-1 |
| [37] |
孙志萍, 秦琳, 刘耀斌, 等. 水稻对富营养化水体生态修复效应及其研究进展[J]. 中国水稻科学, 2018, 32(5): 509-518. SUN Zhiping, QIN Lin, LIU Yao-bin, et al. Advances in restoration effects of rice growing on eutrophic water[J]. Chinese Journal of Rice Science, 2018, 32(5): 509-518. |