 2020, Vol. 39
2020, Vol. 39



CH4是仅次于CO2的第二大温室气体,稻田是CH4排放的重要来源,其排放占我国农业生产CH4总排放的17.83%[1]。水稻品种间CH4排放具有差异性,原因是水稻植株特性的不同会影响CH4产生、氧化、排放的全过程[2]。稻田CH4排放受植株的通气组织[3]、地上部形态特征(分蘖数、株高、叶面积)[4-5]、根系形态特征(根体积与根孔隙度)[5-6]、当季光合产物[7-8]、生物量[9-10]的影响,也受植株根的生理特性影响[11]。同时,稻田CH4排放也是产甲烷菌产生CH4,然后经过甲烷氧化菌氧化以及传输后的净效应[12]。土壤酶是一种蛋白质,具有生物活性,能够参与土壤中的各种生化反应过程且反映其方向和强度[13]。土壤酶也能体现土壤微生物的活性,与微生物的物种丰富度和数量有关。土壤过氧化氢酶(Catalase)作为一种氧化还原酶,能催化过氧化氢的分解,并且将土壤内的有毒物质快速分解成对水稻无害的小分子物质,释放出氧气,可以有效控制土壤氧化的强度和方向[14]。土壤脲酶(Urease)是一种催化尿素分解的水解酶,在土壤中分布较广,数量相对较多。土壤中的氮元素转化与脲酶关系密切,且土壤中的微生物也与脲酶相互作用[15]。土壤中的微生物群落是影响稻田土壤产生CH4的重要因素,对CH4的产生、氧化及排放具有综合影响。研究种植不同水稻品种的条件下两种酶的活性,进而反映出土壤甲烷功能菌的活性,间接揭示其对CH4排放的影响具有重要意义。
减少稻田CH4排放的措施包括低CH4排放品种选择[16]、CH4抑制剂肥料[17]、节水灌溉[18]等,都有较多研究,不论哪种减排技术或者措施,必须是水稻种植主体愿意接受,并且自觉应用才会产生实际作用,种植低排高产的优质水稻品种是容易得到推广应用的措施之一。历年来很多学者围绕不同水稻品种的植株特性与CH4排放的影响等展开研究,试图解释其排放规律,但对于根际土壤酶与CH4排放的研究相对较少。本研究是在肖志祥等[19]研究基础上,进一步监测了双季稻6个品种的CH4排放规律,分析了6个品种全生育期不同过氧化氢酶、脲酶活性的特征,探讨了两种酶与水稻CH4排放量之间的相关性,有助于进一步明确高产水稻品种低CH4排放的机理。
1 材料与方法 1.1 试验地概况试验地点位于浏阳市沿溪镇(113.49°E,28.18°N),为南方典型的双季水稻生产区。试验田土壤为壤土,为第四纪红壤发育的红黄泥水稻土,土壤pH为5.95、全氮3.09 g·kg-1、碱解氮142.89 mg·kg-1、全磷0.67 g· kg-1、速效磷53.77 mg·kg-1、全钾7.52 g·kg-1、速效钾98.02 mg·kg-1、有机质含量34.21 g·kg-1。试验点2019年的日平均降雨量与温度如图 1所示。

|
图 1 2019年试验点气温与降雨量日变化 Figure 1 Variation of daily air temperature and precipitation of 2019 at the research station |
供试早稻品种:株两优819、陆两优996、湘早籼24号;晚稻品种:Y两优1号、泰优390、湘晚籼17号。早稻播种日期为2019年3月25日,移栽:4月29日,收获:7月21日;晚稻播种日期为6月22日,移栽:8月3日,收获:10月31日。早稻、晚稻试验为随机区组设计,3个品种处理,3次重复,共9个小区,小区面积36 m2(4 m×9 m)。所有试验小区的管理与当地传统田间管理方式一致,氮肥分基肥、蘖肥、穗肥3次施入,比例为5:3:2,早稻施氮量(折合为纯氮)120 kg·hm-2,晚稻施氮量150 kg·hm-2。磷肥做基肥一次性施入。钾肥分基肥、蘖肥两次施入,比例为1:1。早稻和晚稻均施用过磷酸钙(P2O5 12%)750 kg·hm-2、氯化钾(K2O 60%)201 kg·hm-2。
1.3 观测指标与方法 1.3.1 温室气体取样与测定通过静态暗箱法收集CH4气体样品,在湖南省农业科学院环境研究所使用Agilent 7890A气相色谱仪分析气体样品,CH4采用FID检测器,检测温度为180 ℃,柱温70 ℃,载气流速40 mL·min-1,由国家标准物质中心提供标准气体。采样箱为长方体形状,规格为0.5 m×0.5 m×1.2 m,底座规格为0.5 m×0.5 m×1.2 m,为达到更好的密闭及保温效果,用泡沫板覆盖且用胶带缠封气体箱四周,为充分混合箱内气体,在其内部装一个12 V小风扇,用温度传感器记录采样时箱内温度变化情况。水稻移栽前,将采样箱底座齐平土层表面插入每个小区内。秧苗移栽扎根立苗后,选择晴朗天气(9:00—11:00)进行取样,约7 d取一次气体样品,早稻于5月13日、晚稻于8月7日开始取气,直至水稻成熟。采样箱扣进底座时,注意要向底座凹槽注水以达到箱内密封状态,并及时用带有三通阀的60 mL针筒与取气箱的三通阀连接,每间隔10 min取一次样,连续取样4次,每次抽样体积为45 mL,并及时注入真空瓶中,同时记录箱体内温度。稻田温室气体排放通量以及季节累积排放量计算公式参考杨丹等[20]研究。
全球增温潜势(Global warming potential,GWP)是将CH4排放折算为CO2当量,其系数是25,温室气体排放强度(Green house gas intensity,GHGI)是GWP与实际产量的比值。
1.3.2 土壤样品采集与测定在水稻分蘖期、孕穗期、齐穗期、乳熟期、成熟期5个时期采用五点取样法取样,将水稻植株连同根系土壤一起用铁铲挖出,抖落掉根系外围土层,用无菌手套采取距根际较近的土层(约1~2 cm)作为土样。在自然条件下晾干,然后按照所需筛目数进行过筛,用于酶活性的检测。脲酶采用靛酚蓝比色法,脲酶活性以24 h后1 g土壤中NH3-N的毫克数表示。过氧化氢酶采用KMnO4滴定法,由每克干土20 min内用0.02 mol·L-1高锰酸钾滴定酶促反应前后过氧化氢所需毫升数之间的差值求出酶促反应分解过氧化氢的量,以此表示过氧化氢酶活性。
1.4 数据统计分析采用SPSS 20.0软件进行统计分析,并作相关显著性检验(P < 0.05)及回归分析;采用Excel 2003整理数据以及作图。图表中数据为平均值±标准差。
2 结果与分析 2.1 双季稻不同品种CH4排放差异 2.1.1 双季稻品种CH4排放通量动态变化早稻CH4排放通量动态变化如图 2所示,水稻CH4排放有两个波峰,第一次是在分蘖期,第二次是在孕穗期,孕穗期后,CH4排放通量开始缓慢下降并在乳熟期时达到最低,之后变化不显著。在5月13日至6月3日,湘早籼24号的CH4排放量始终大于其他两个品种。晚稻3个品种的CH4排放动态变化基本上一致,具有两个明显的排放高峰值,第一次稻田CH4排放的最高峰值出现在水稻移栽后的短时间内(10~ 15 d),而后CH4的排放通量迅速下降,并维持在较低水平,第二次CH4排放峰值在晚稻生长的后期(9月4日),之后排放没有明显相同规律,但都维持在一个较低水平。

|
图 2 早稻晚稻不同品种CH4排放通量变化 Figure 2 Variation of CH4 emission flux in different early rice and late rice varieties |
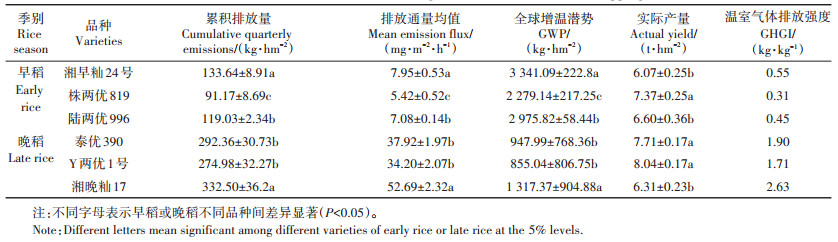
不同双季稻品种间CH4排放差异显著(表 1)。早稻品种的CH4累积排放量介于91.17~133.64 kg·hm-2,排放量最低是株两优819,为91.17 kg·hm-2,分别较湘早籼24号、陆两优996的排放通量低31.78%、23.41%,与两者差异显著(P < 0.05)。早稻株两优819的全球增温潜势(GWP)较湘早籼24号、陆两优996分别低31.78%、23.41%;早稻温室气体排放强度(GHGI)介于0.31~0.55 kg·kg-1,株两优819最低。晚稻品种的CH4累积排放量明显高于早稻,介于274.98~332.50 kg·hm-2,以Y两优1号最低,湘晚籼17号最高,泰优390与Y两优1号差异不显著(P>0.05);Y两优1号分别较湘晚籼17号和泰优390的排放通量偏低35.09%和9.81%(P < 0.05)。晚稻Y两优1号的GWP较湘晚籼17号、泰优390,分别低17.31%、5.94%;晚稻的GHGI介于1.71~2.63 kg·kg-1,Y两优1号最低。
|
|
表 1 双季稻CH4累积排放量和温室效应 Table 1 Methane cumulative emissions and greenhouse effect of double cropping rice |
早稻3个品种,根际土壤过氧化氢酶活性大小范围为0~2.88 mL KMnO4·g-1·min-1,不同时期过氧化氢酶活性大小均为:孕穗期>分蘖期>乳熟期>齐穗期>成熟期(图 3);其中在分蘖期、孕穗期、成熟期,3者差异不显著(P > 0.05)。在水稻孕穗之前,过氧化氢酶活性大小为:湘早籼24>株两优819>陆两优996。在齐穗期和乳熟期,陆两优996的过氧化氢酶活性明显高于另外两个早稻品种(P < 0.05),而株两优819和湘早籼24号的过氧化氢酶活性差异不显著。

|
不同字母表示不同品种间差异显著(P < 0.05)。下同 Different letters show significant difference among different varieties at 0.05 level. The same below 图 3 双季稻不同生育期过氧化氢酶活性 Figure 3 Catalase activity in different growth stages of double cropping rice |
晚稻品种的根际土壤过氧化氢酶介于4.50~5.84 mL KMnO4·g-1·min-1,总体上呈先降低后升高的趋势(图 3)。在晚稻分蘖期、孕穗期时,各品种过氧化氢酶活性大小为:泰优390>Y两优1号>湘晚籼17号,且分蘖期3者具有明显差异(P < 0.05),孕穗期,泰优390明显高于另外两个品种,Y两优1号和湘晚籼17号的过氧化氢酶活性没有明显差异;在齐穗期,3个品种过氧化氢酶活性大小为:湘晚籼17号>泰优390>Y两优1号,3者具有明显差异(P < 0.05);而在乳熟期,3个品种酶活性大小刚好与齐穗期相反,此时,Y两优1号的过氧化氢酶活性最大;在成熟期,泰优390的过氧化氢酶活性最大,湘晚籼17号的最小。
2.2.2 双季稻品种脲酶活性由图 4可知,湘早籼24号和陆两优996的脲酶活性在整个生育期呈先降低后升高然后再降低,在成熟期脲酶活性最低;株两优819有细微差异,呈先降低,后在成熟期有所升高。在水稻分蘖期,陆两优996的脲酶活性明显大于湘早籼24号和株两优819(P < 0.05);在孕穗期和乳熟期,3个品种脲酶活性大小均为:陆两优996>株两优819>湘早籼24号,陆两优996与株两优819差异不显著;在齐穗期,湘早籼24号和陆两优996的脲酶活性明显大于株两优996;在成熟期,株两优819的脲酶活性明显大于陆两优996和湘早籼24号,但湘早籼24号和陆两优996差异不显著。

|
图 4 双季稻不同生育期脲酶活性 Figure 4 Urease activity in different growth stages of double cropping rice |
晚稻Y两优1号根际土壤脲酶活性随生育期而降低,泰优390和湘晚籼17号脲酶活性在不同生育期变化趋于一致,基本上呈“S”形变化,先降低后有所升高。在水稻分蘖期,3个品种的脲酶活性具有明显差异,Y两优1号的脲酶活性最高,其次是泰优390,最低的是湘晚籼17号;在孕穗期、齐穗期3个品种的脲酶活性大小为:Y两优1号>泰优390>湘晚籼17号,Y两优1号与泰优390的脲酶活性差异不显著,而两者的脲酶活性与湘晚籼17号具有明显差异;在乳熟期,3个品种的脲酶活性没有明显差异;在成熟期,泰优390的脲酶活性有所升高,并且明显高于湘晚籼17号,Y两优1号与泰优390的脲酶活性差异不显著。
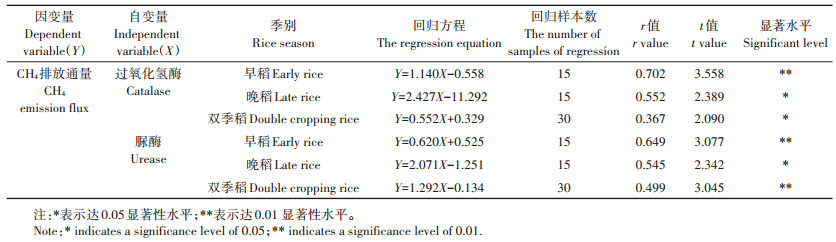
2.3 双季稻品种CH4排放通量与土壤酶的相关分析将水稻品种在分蘖期、孕穗期、齐穗期、乳熟期、成熟期5个时期的土壤酶活性与对应5个时期的CH4排放通量作回归分析,由表 2可知,双季稻CH4排放通量(Y)与脲酶活性(X)呈极显著正相关(P < 0.01),与过氧化氢酶活性(X)呈显著正相关(P < 0.05);早稻CH4排放通量与早稻过氧化氢酶、脲酶的活性呈极显著正相关(P < 0.01),晚稻CH4排放通量与晚稻过氧化氢酶、脲酶的活性呈显著正相关(P < 0.05)。早稻CH4排放通量与两种酶的相关性比晚稻好,其与早稻过氧化氢酶活性相关系数最高(r=0.702**)。本试验研究结果表明,双季稻田土壤过氧化氢酶、脲酶活性越高,其CH4排放通量越高,其中脲酶活性更显著影响CH4排放通量。
|
|
表 2 双季稻CH4排放通量与土壤酶活性的回归系数及方差分析 Table 2 The regression coefficient CH4 emission flux and soil enzyme activity in double cropping rice |
本试验6个品种CH4排放差异显著,各品种CH4累积排放量大小与肖志祥等[19]研究相同,排放趋势也有相似规律,排放峰值出现在生育前期,且早稻和晚稻CH4排放趋势相似。本研究发现,早稻晚稻的3个品种CH4排放通量不同,在不同生育期基本呈现一致的变化规律,其有两个波峰,变化趋势为先增后减。这一研究结果与Wang等[21]和孙会峰等[22]的研究结果接近,分蘖盛期水稻生长繁茂,分蘖数量多,CH4排放通道多,有利于CH4排放。此外,水稻在晒田时期CH4排放通量最低。Li等[23]也研究发现水稻在中期晒田时CH4排放通量最低,甚至接近0排放。晒田中期,稻田土壤表面水分急剧减少,土壤通透性增加,使得大量氧气进入土壤,土壤氧化CH4的能力得到了提高,抑制产CH4菌的活性,从而导致CH4排放的减少[24]。晒田后复水,施入穗肥,稻田的有机质增加,造成了水稻排放通量的第二个峰值。晚稻CH4排放通量明显高于早稻,且晚稻CH4排放峰值出现时间也较早。原因在于晚稻较早稻生长期气温高且降雨量少,田间需不断回水促进生长,为产CH4菌营造了有利的生长环境,且早晚稻植株长势不一致,水稻是喜温作物,晚稻光温资源较早稻丰沛,植株生长较快,通气组织发达,促进稻田CH4排放。
3.2 双季稻品种温室效应的差异本研究中,早稻3个品种CH4累积排放量具有显著差异,表现为籼系杂交稻明显低于籼系常规稻;晚稻3个品种中,杂交籼稻品种累积排放量差异不显著,但都明显低于常规稻[22, 25-26]。中国水稻种植区晚稻CH4累积排放量高于早稻[27],本试验3个品种晚稻CH4累积排放量是早稻3个品种的1.06~2.65倍。不同水稻品种增温潜势中,早稻以株两优819最低,明显较早稻其他品种低23.41%~31.78%,晚稻以Y两优1号最低,较晚稻其他品种低5.94%~17.31%。温室气体排放强度均与水稻产量相关,本试验GHGI均与GWP大小特征表现一致。同样地,所有供试品种产量具有差异,早稻最高产量较最低产量高51.11 kg,晚稻最高产量较最低产量高96.43 kg。在本试验研究结果中,株两优819与Y两优1号具有高产低排的特征。
3.3 双季稻品种过氧化氢酶与脲酶对CH4排放的影响水稻的根际土壤环境是影响植株CH4产生与氧化的重要因素之一,土壤酶作为评价土壤肥力和质量的重要指标[28],其主要来源是微生物和植物根系。土壤酶与温室气体的产生息息相关,一方面,酶的催化作用会影响土壤微生物的活性,加速根系生长,刺激根系分泌物产生,同时根际微区活动旺盛也会增加土壤酶的活性,根系与微生物的协同作用促进了CH4的排放[29];另一方面,土壤酶参与有机质的矿化、土壤氮的氧化还原等生化反应[28],有研究表明,过氧化氢酶与有机质、全氮、速效氮具有正效应[30],脲酶活性增加,会加速尿素分解,增加供氮水平,参与土壤氮素循环[31],两种酶活性越高,为产甲烷菌提供的底物越多,越有利于CH4排放。
本研究结果表明,晚稻品种的过氧化氢酶活性均高于早稻品种,早稻品种在全生育时期内具有明显一致的变化特征:分蘖期酶活性不断升高,孕穗期后均不断下降,晚稻各个品种全生育期没有明显一致的变化趋势,但所有品种在孕穗期都表现为下降趋势。相关研究表明,土壤过氧化氢酶活性在植物生育前期较高,分蘖结束后明显降低,拔节期后酶活性趋于稳定[18],这可能是由于土壤总生物量和生物呼吸量影响其活性[32]。有研究表明[33-34],土壤酶的数量越高,活性越强,其土壤中的微生物数量较多,繁殖速度快。根际土壤脲酶活性表现为晚稻3个品种脲酶活性差异性比早稻3个品种更大,并且早晚稻脲酶活性齐穗前大于齐穗后,早晚稻成熟期脲酶活性低于乳熟期。早稻脲酶活性大小介于0.37~4.34 mg·g-1·d-1,晚稻脲酶活性介于0.77~3.90 mg·g-1·d-1。尿素是有机态氮肥,施入土壤后,一部分以分子态溶于土壤溶液中,通过氢键作用被土壤吸附,其他大部分在脲酶的作用下水解成NH3和CO2,然后以NH4+被植物吸收和土壤胶体吸附[35]。陆两优996和Y两优1号的脲酶活性高,其对尿素分解的催化作用强,使两者吸氮能力强,从而水稻产量也是最高的,且脲酶活性与双季稻CH4排放具有极显著正相关关系(r=0.499**)。
4 结论(1)不同双季稻品种的CH4排放高峰期主要是分蘖期和孕穗期,最高排放峰值出现在分蘖期,在乳熟期后水稻的CH4排放较低。早稻品种CH4累积排放量、温室效应表现为:湘早籼24号>陆两优996>株两优819;晚稻品种CH4累积排放量、温室效应表现为:湘晚籼17号>泰优390>Y两优1号。CH4排放最低的株两优819和Y两优1号产量最高。
(2)在CH4排放高峰期,双季稻品种过氧化氢酶、脲酶活性具有相同的大小规律,两种酶活性均为前期高于生育后期,且早稻品种中,株两优819的过氧化氢酶活性最低,脲酶活性仅次于陆两优996,晚稻品种中,Y两优1号的过氧化氢酶活性仅次于泰优390,脲酶活性最高。
(3)水稻品种CH4排放通量与土壤过氧化氢酶、脲酶活性具有正线性相关,显著性水平分别达P= 0.05、P=0.01。这一结论可为今后进一步探究高产低排品种机理提供参考。
| [1] |
国家发展和改革委员会应对国家气候变化司. 中华人民共和国气候变化第二次国家信息通报[M]. 北京: 中国经济出版社, 2013: 28-30. National Climate Change Division of the National Development and Reform Commission. Second national communication of the People's Republic of China on climate change[M]. Beijing: China Economic Press, 2013: 28-30. |
| [2] |
江瑜, 管大海, 张卫建. 水稻植株特性对稻田甲烷排放的影响及其机制的研究进展[J]. 中国生态农业学报, 2018, 26(2): 175-181. JIANG Yu, GUAN Da-hai, ZHANG Wei-jian. The effect of rice plant traits on methane emissions from paddy fields:A review[J]. Chinese Journal of Eco-Agriculture, 2018, 26(2): 175-181. |
| [3] |
李奕林. 水稻根系通气组织与根系泌氧及根际硝化作用的关系[J]. 生态学报, 2012, 32(7): 2066-2074. LI Yi-lin. Relationship among rice root aerechyma, root radial oxygen loss and rhizosphere nitrification[J]. Acta Ecologica Sinica, 2012, 32(7): 2066-2074. |
| [4] |
Gogoi N, Baruah K K, Gupta P K. Selection of rice genotypes for lower methane emission[J]. Agronomy for Sustainable Development, 2008, 28(2): 181-186. DOI:10.1051/agro:2008005 |
| [5] |
Das K, Baruah K K. Methane emission associated with anatomical and morphophysiological characteristics of rice(Oryza sativa) plant[J]. Physiologia Plantarum, 2008, 134(2): 303-312. |
| [6] |
Singh S, Singh J S, Kashyap A K. Methane flux from irrigated rice fields in relation to crop growth and N-fertilization[J]. Soil Biology and Biochemistry, 1999, 31(9): 1219-1228. DOI:10.1016/S0038-0717(99)00027-9 |
| [7] |
Tokida T, Adachi M, Cheng W, et al. Methane and soil CO2 production from current-season photosynthates in a rice paddy exposed to elevated CO2 concentration and soil temperature[J]. Global Change Biology, 2011, 17(11): 3327-3337. DOI:10.1111/j.1365-2486.2011.02475.x |
| [8] |
Yuan Q, Pump J, Conrad R. Partitioning of CH4 and CO2 production originating from rice straw, soil and root organic carbon in rice microcosms[J]. PLoS One, 2012, 7(11): e49073. DOI:10.1371/journal.pone.0049073 |
| [9] |
Huang Y, Sass R, Fisher F. Methane emission from Texas rice paddy soils. 2. Seasonal contribution of rice biomass production to CH4 emission[J]. Global Change Biology, 1997, 3(6): 491-500. DOI:10.1046/j.1365-2486.1997.00106.x |
| [10] |
Van G K J, Van K C, Hungate B A. Increased greenhouse-gas intensity of rice production under future atmospheric conditions[J]. Nature Climate Change, 2013, 3(3): 288-291. DOI:10.1038/nclimate1712 |
| [11] |
Aulakh M S, Wassmann R, Bueno C, et al. Impact of root exudates of different cultivars and plant development stages of rice(Oryza sativa L.)on methane production in a paddy soil[J]. Plant and Soil, 2001, 230(1): 77-86. DOI:10.1023/A:1004817212321 |
| [12] |
夏龙龙, 颜晓元, 蔡祖聪. 我国农田土壤温室气体减排和有机碳固定的研究进展及展望[J]. 农业环境科学学报, 2020, 39(4): 834-841. XIA Long-long, YAN Xiao-yuan, CAI Zu-cong. Research progress and prospect of greenhouse gas mitigation and soil carbon sequestration in croplands of China[J]. Journal of Agro-Environment Science, 2020, 39(4): 834-841. |
| [13] |
Yang Z, Liu S, Zheng D, et al. Effects of cadium, zinc and lead on soil enzyme activityity[J]. Journal of Environmental Sciences, 2006, 18(6): 1135-1141. DOI:10.1016/S1001-0742(06)60051-X |
| [14] |
桓明辉, 李杨, 刘晓辉, 等. 秸秆生物菌肥对保护地土壤微生物和土壤酶活性的影响[J]. 山东农业科学, 2013, 45(1): 107-109, 116. HUAN Ming-hui, LI Yang, LIU Xiao-hui, et al. Effects of straw and bio-bacterial manure on microorganism and enzyme activity in protected vegetable soil[J]. Shandong Agricultural Sciences, 2013, 45(1): 107-109, 116. |
| [15] |
马维娜, 杨京平, 汪华. 不同水分模式分次施氮对水稻根际土壤微生物生态效应的影响[J]. 浙江大学学报(农业与生命科学版), 2007, 33(2): 184-189. MA Wei-na, YANG Jing-ping, WANG Hua. Ecological effect of rice rhizosphere microbes under water regimes and nitrogen fertilizer with split application in paddy field[J]. Journal of Zhejiang University(Agriculture and Life Sciences), 2007, 33(2): 184-189. |
| [16] |
黄农荣, 梁开明, 钟旭华, 等. 南方低甲烷排放的高产水稻品种筛选与评价[J]. 农业环境科学学报, 2018, 37(12): 2854-2863. HUANG Nong-rong, LIANG Kai-ming, ZHONG Xu -hua, et al. Screening for and evaluation of rice(Oryza sativa)varieties with low methane emission and high yield in South China[J]. Journal of AgroEnvironment Science, 2018, 37(12): 2854-2863. |
| [17] |
周旋, 吴良欢, 戴锋, 等. 生化抑制剂组合与施肥模式对黄泥田稻季CH4和N2O排放的影响[J]. 生态与农村环境学报, 2018, 34(12): 1122-1130. ZHOU Xuan, WU Liang-huan, DAI Feng, et al. Effects of combined biochemical inhibitors and fertilization models on CH4 and N2O emission from yellow clayey field during rice growth season[J]. Journal of Ecology and Rural Environment, 2018, 34(12): 1122-1130. |
| [18] |
李健陵, 李玉娥, 周守华, 等. 节水灌溉、树脂包膜尿素和脲酶/硝化抑制剂对双季稻温室气体减排的协同作用[J]. 中国农业科学, 2016, 49(20): 3958-3967. LI Jian-ling, LI Yu-e, ZHOU Shou-hua, et al. Synergistic effects of water-saving irrigation, polymer-coated nitrogen fertilizer and urease/nitrification inhibitor on mitigation of greenhouse gas emissions from the double rice cropping system[J]. Scientia Agricultura Sinica, 2016, 49(20): 3958-3967. |
| [19] |
肖志祥, 傅志强, 徐华勤, 等. 双季稻品种根际特征与甲烷排放差异及其关系[J]. 环境科学, 2019, 40(2): 904-914. XIAO Zhi-xiang, FU Zhi-qiang, XU Hua-qin, et al. Differences and relationship between rhizosphere characteristics and methane emissions of double-cropping rice variety[J]. Environmental Science, 2019, 40(2): 904-914. |
| [20] |
杨丹, 叶祝弘, 肖珣, 等. 化肥减量配施有机肥对早稻田温室气体排放的影响[J]. 农业环境科学学报, 2018, 37(11): 2443-2450. YANG Dan, YE Zhu-hong, XIAO Xun, et al. Effects of chemical fertilizer reduction and organic fertilizer use on the greenhouse gas emissions of early rice fields[J]. Journal of Agro-Environment Science, 2018, 37(11): 2443-2450. |
| [21] |
Wang J Y, Zhang X L, Xiong Z Q, et al. Methane emissions from a rice agroecosystem in South China:Effects of water regime, straw incorporation and nitrogen fertilizer[J]. Nutrient Cycling in Agroecosystems, 2012, 93(1): 103-112. DOI:10.1007/s10705-012-9503-3 |
| [22] |
孙会峰, 周胜, 陈桂发, 等. 水稻品种对稻田CH4和N2O排放的影响[J]. 农业环境科学学报, 2015, 34(8): 1595-1602. SUN Hui-feng, ZHOU Sheng, CHEN Gui-fa, et al. Effects of rice cultivars on CH4 and N2O emissions from rice fields[J]. Journal of AgroEnvironment Science, 2015, 34(8): 1595-1602. |
| [23] |
Li X, Yuan W, Xu H, et al. Effect of timing and duration of midseason aeration on CH4 and N2O emissions from irrigated lowland rice paddies in China[J]. Nutrient Cycling in Agroecosystems, 2011, 91(3): 293-305. DOI:10.1007/s10705-011-9462-0 |
| [24] |
Jia Z J, Cai Z C, Xu H, et al. Effect of rice plants on CH4 production, transport, oxidation and emission in rice paddy soil[J]. Plant and Soil, 2001, 230(2): 211-221. DOI:10.1023/A:1010366631538 |
| [25] |
傅志强, 黄璜, 何保良, 等. 水稻植株通气系统与稻田CH4排放相关性研究[J]. 作物学报, 2007, 33(9): 1458-1467. FU Zhi-qiang, HUANG Huang, HE Bao-liang, et al. Correlation between rice plant aerenchyma system and methane emission from paddy field[J]. Acta Agronomica Sinica, 2007, 33(9): 1458-1467. |
| [26] |
傅志强, 黄璜, 朱华武, 等. 水稻CH4和N2O的排放及其与植株特性的相关性[J]. 湖南农业大学学报(自然科学版), 2011, 37(4): 356-360. FU Zhi-qiang, HUANG Huang, ZHU Hua-wu, et al. Relativity between CH4 and N2O emission and rice plant characteristics[J]. Journal of Hunan Agricultural University(Natural Sciences), 2011, 37(4): 356-360. |
| [27] |
魏海苹, 孙文娟, 黄耀. 中国稻田甲烷排放及其影响因素的统计分析[J]. 中国农业科学, 2012, 45(17): 3531-3540. WEI Hai-ping, SUN Wen-juan, HUANG Yao. Statistical analysis of methane emission from rice fields in China and the driving factors[J]. Scientia Agricultura Sinica, 2012, 45(17): 3531-3540. |
| [28] |
关松荫, 郑洪元. 土壤酶及其研究法[M]. 北京: 中国农业出版社, 1986: 14-18. GUAN Song-yin, ZHENG Hong-yuan. Soil enzymes and their research methods[M]. Beijing: China Agriculture Press, 1986: 14-18. |
| [29] |
甄丽莎, 谷洁, 高华, 等. 秸秆还田与施肥对土壤酶活性和作物产量的影响[J]. 西北植物学报, 2012, 32(9): 1811-1818. ZHEN Li-sha, GU Jie, GAO Hua, et al. Effect of straws, manure and chemical fertilizer on soil properties and crop yields[J]. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(9): 1811-1818. |
| [30] |
陈一民, 焦晓光, 王贵强, 等. 空间移位下农田黑土过氧化氢酶与土壤养分的相关性研究[J]. 黑龙江大学工程学报, 2012, 3(4): 46-50. CHEN Yi-min, JIAO Xiao-guang, WANG Gui-qiang, et al. Relationship between catalase activities and soil nutrients for farmland phaeozem based on soil removal method[J]. Journal of Engineering of Heilongjiang University, 2012, 3(4): 46-50. |
| [31] |
邓欧平, 李翰, 熊雷, 等. 秸秆、猪粪混施对麦田根际土壤过氧化氢酶与蔗糖酶活性的影响[J]. 土壤, 2018, 50(1): 86-92. DENG Ou-ping, LI Han, XIONG Lei, et al. Effect of mixed application of rice-wheat straws and pig manure on soil enzyme activities in wheat-growth field[J]. Soils, 2018, 50(1): 86-92. |
| [32] |
杨鹏鸣, 周俊国. 不同肥料对土壤蔗糖酶和过氧化氢酶活性的影响[J]. 广东农业科学, 2011, 38(11): 78-80. YANG Peng-ming, ZHOU Jun-guo. Effects of different fertilizer on soil invertase and catalase activity[J]. Guangdong Agricultural Sciences, 2011, 38(11): 78-80. |
| [33] |
马志良, 赵文强, 刘美, 等. 高寒灌丛生长季土壤转化酶与脲酶活性对增温和植物去除的响应[J]. 应用生态学报, 2018, 29(7): 2211-2216. MA Zhi-liang, ZHAO Wen-qiang, LIU Mei, et al. Responses of soil invertase and urease activities to warming and plant removal during the growing season in an alpine scrub ecosystem[J]. Chinese Journal of Applied Ecology, 2018, 29(7): 2211-2216. |
| [34] |
张知晓, 泽桑梓, 户连荣, 等. 土壤脲酶活性调控因素和脲酶活性细菌系统发育研究[J]. 西部林业科学, 2018, 47(1): 65-73. ZHANG Zhi-xiao, ZE Sang-zi, HU Lian-rong, et al. Regulatory factors of soil urease activity and phylogenetic analysis of urease bacteria[J]. Journal of West China Forestry Science, 2018, 47(1): 65-73. |
| [35] |
张志勇, 于旭昊, 熊淑萍, 等. 耕作方式与氮肥减施对黄褐土麦田土壤酶活性及温室气体排放的影响[J]. 农业环境科学学报, 2020, 39(2): 418-428. ZHANG Zhi-yong, YU Xu-hao, XIONG Shu-ping, et al. Effects of tillage methods and nitrogen fertilizer reduction on soil enzyme activities and greenhouse gas emissions of wheat yellow cinnamon soil[J]. Journal of Agro-Environment Science, 2020, 39(2): 418-428. |