 2021, Vol. 40
2021, Vol. 40



2. 湖南省湘潭市农业科学研究所, 湖南 湘潭 411134
2. Xiangtan Agricultural Science Research Institute, Xiangtan 411134, China
我国大约有65% 人口以稻米为主食,稻米的营养成分与人类健康密切相关[1]。当水稻生长在Cd污染土壤中时,Cd能在其根、茎、叶以及籽粒中大量积累,不仅影响水稻的生长发育,而且还严重影响稻米品质,并经食物链危及人类和动物的健康[2]。全世界大约有2.35×1012 m2的耕地被重金属污染[3],在有害金属中,Cd是一种流动性强且毒性大的重金属[4-5];在我国南方酸性土壤中,即使稻田土壤Cd含量在0.3 mg·kg-1以下,也有可能生产出Cd超标的稻米[6-7]。因此,有效控制稻米Cd积累的技术措施对于中轻度Cd污染农田的安全利用具有重要意义。
苹果酸(Malic acid,MA)是三羧酸循环(Tricarboxylic acid cycle,简称TCA循环)及其支路乙醛酸循环代谢过程中的重要中间产物[8],可以迅速通过生物膜,进入线粒体内直接参与能量代谢,对于NADH的转移和ATP的合成发挥着重要的作用[9]。为了保证细胞各种代谢活动对苹果酸的需求,水稻和十字花科植物体内进化出多种苹果酸酶(NADP-ME)基因家族[10-12]。当植物受到重金属、干旱、盐碱、高温等非生物胁迫时,NADP-ME基因的表达水平和细胞中的苹果酸积累量会显著增加[10-11, 13]。外源添加苹果酸能显著提高植物生长量,增加净光合速率,减少H2O2的积累,增强根系活性,抑制根系对Cd的吸收和转运等,进而减轻Cd的毒害作用[14-18]。水稻开花期叶面喷施苹果酸,对水稻籽粒、穗轴、穗颈和旗叶中的Cd含量能产生显著的抑制作用,对Mn、Zn等营养元素的转运也有显著的影响[19]。这些研究结果表明,苹果酸能通过调控营养元素平衡、消除氧化损伤等多种方式缓解Cd对水稻产生的生理毒害作用。
植物抗逆指标有多种,抗氧化酶活性、可溶性蛋白、可溶性氨基酸、氨基酸含量都和酶的活性有关,Cd胁迫和其他非生物胁迫都会引起抗氧化酶活性的变化。在Cd等重金属胁迫环境中,水稻细胞经常通过合成大量的金属硫蛋白、谷胱甘肽(GSH)、植物螯合素(PCs)来缓解Cd的毒害[20]。氨基酸是蛋白质的基本成分,对维持细胞的正常代谢非常重要。水稻籽粒中的蛋白质和氨基酸主要分布在胚和糊粉层中[21],能与Cd形成螯合物使其失去生物毒性。谷氨酸、丙氨酸、苯丙氨酸、甘氨酸和苏氨酸5种氨基酸与水稻籽粒中Cd含量高度线性相关;谷氨酸的合成能力越强,籽粒中富集Cd的能力越强[22]。增施钙镁磷肥既能抑制Cd向稻米中的转运,又能促进稻米中氨基酸的合成[23]。这些结果说明,氨基酸代谢与稻米的Cd积累特性密切相关。因此,本研究以氨基酸含量为主要指标,来研究氨基酸代谢与稻米Cd积累特性及苹果酸降Cd效应的关系。
苹果酸是植物根系和叶片中最丰富的小分子酸,并受植物种类、生长条件等多种因素的影响[9, 24-26]。叶面喷施苹果酸,除了能够抑制稻米中Cd含量外,对稻米的氨基酸含量是否会产生影响,其降Cd效果在水稻品种间是否能够保持稳定,尚未见相关研究报道。本研究以4个水稻品种为材料,采用水稻开花期叶面喷施苹果酸的方法,对成熟期水稻各器官的Cd积累特性和籽粒中的氨基酸含量进行分析,对苹果酸的降Cd效果和机理进行了探讨,旨在为降Cd叶面调理剂的筛选和研发提供参考依据。
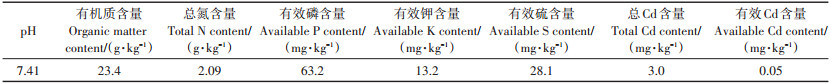
1 材料与方法 1.1 试验材料试验供试土壤为天津市重金属污染农田0~20 cm的表层土,于2019年5—10月在农业农村部环境保护科研监测所日光温室中进行,土壤基本理化性质见表 1。外源喷施的苹果酸(C4H6O5)为分析纯化学试剂。
|
|
表 1 供试土壤基本理化性质 Table 1 The basic physicochemical properties of soil |
根据多年的田间试验和室内试验,筛选出了4个Cd积累特性不同的品种,其中两个是湖南省大面积推广的籼稻品种“T优705”(705)和“湘早籼24号”(X24)[27-28];另外两个是从国家水稻核心资源库提供的核心种质中筛选出的品种“铁杆乌”(TGW)和“辐品36”(F36)[29]。
1.3 样品的处理与采集选取饱满均一的水稻种子在1% 的次氯酸钠溶液中浸泡15 min进行消毒,用去离子水洗净,置于培养皿中保持湿润状态,在28 ℃恒温箱里发芽[28],待胚芽露白后播种于育苗盘中,放在人工智能气候室中,水稻苗长到两叶一心后将育苗盘中的水换成3 L左右1/4 Hoagland营养液,长到四叶一心之后,选取长势均一的幼苗,将根冲洗干净移栽到事先准备好的盆中。水稻扬花期进行叶面喷施处理,处理1喷施去离子水(CK);处理2喷施2次5 mmol·L-1苹果酸(MA1);处理3喷施3次5 mmol·L-1苹果酸(MA2);每次喷施间隔12 h,每盆喷施50 mL,每个处理重复6次。水稻成熟期收取水稻籽粒、穗颈、旗叶、穗节、茎秆等样品进行前处理,用于测定Cd含量、必需元素含量、籽粒氨基酸含量等指标。人工气候室(宁波市科技园区新江南仪器有限公司)条件:白天温度28 ℃,湿度80%,光照16 h,光照强度400 μmol·m-2·s-1;夜间温度20 ℃,湿度70%,8 h黑暗。
1.4 Cd和矿质元素及籽粒氨基酸的测定方法参照Liu等[30]的方法,对样品进行清洗后,用5 mmol·L-1CaCl2浸泡根系15 min后用去离子水洗净并烘干。称取根系与地上部各0.20 g放于消煮管内,加入7 mL硝酸与1 mL双氧水进行消解试验,消解后采用ICP-MS测定样品中Cd、Ca、Fe、Mn、Zn含量。参照Yuan等[22]的方法,称取0.25 g籽粒,加入15 mL 6 mmol·L-1的HCl在110 ℃下水解22 h,用去离子水定容至50 mL,吸取1 mL于50 ℃水浴中氮气干燥,最后用柠檬酸盐缓冲溶液重新溶解。溶解后采用高效液相色谱系统(Agilent Technologies,Palo Alto,CA)进行氨基酸含量分析。
1.5 数据统计及分析采用Microsoft Excel进行相关数据的计算、统计与处理。Cd含量和必需元素含量用SPSS 17.0进行二因子随机设计方差分析;氨基酸含量用SPSS 17.0进行单因子随机设计方差分析,新复极差法(Duncan)作多重比较(同一器官的品种和苹果酸浓度为12个处理进行多重比较)、差异显著性检验,并用Origin 8.6作图。
转移系数为相邻器官间Cd含量的比值[31]:
转移系数(TFa/b)=a器官Cd含量/b器官Cd含量
2 结果与分析 2.1 苹果酸对稻米Cd积累特性及转移系数的影响Cd含量在不同品种之间以及水稻各器官间存在明显的差异。穗节中的Cd含量最高,穗颈次之,籽粒和旗叶最低;穗节中的Cd含量是穗颈和旗叶Cd含量的2~10倍(图 1A、图 1B)。虽然4个品种的开花期和生育期非常接近,但705和X24的穗节、穗颈、茎秆中的Cd含量明显高于F36和TGW,说明品种的遗传背景对各器官Cd积累能力有显著影响。

|
不同小写字母表示处理间差异达到5%显著水平。下同 Different letters indicate significant differences among treatments at 5% level. The same below 图 1 水稻开花期叶面喷施苹果酸对水稻各器官Cd含量以及相邻器官Cd转移系数的影响 Figure 1 Effects of foliar application with malic acid at anthesis stage on Cd content in rice organs and transfer factors of Cd between jointed organs in rice plants |
虽然各器官的Cd含量在品种间有明显差异,但喷施苹果酸后产生的降Cd效应更为显著,为了消除基因型及环境对苹果酸效应的影响,显示苹果酸浓度对Cd含量的影响,将4个品种进行了平均值的比较。结果显示,籽粒、穗颈、旗叶、穗节中Cd含量的下降幅度随着苹果酸喷施次数的增加而加大。与CK相比,喷施2次和3次苹果酸后,4个品种籽粒中的Cd平均含量从0.26 mg·kg-1分别下降到0.14 mg·kg-1和0.09 mg·kg-1(图 1C),降幅分别为46.15% 和65.38%(图 1D);穗颈中的Cd平均含量从0.51 mg·kg-1分别下降到0.31 mg·kg-1和0.18 mg·kg-1,降幅分别为39.22%和64.71%;穗节中的Cd平均含量从1.60 mg·kg-1分别下降到1.04 mg·kg-1和0.69 mg·kg-1,降幅分别为35.00% 和56.88%;旗叶中的Cd平均含量从0.18 mg·kg-1分别下降到0.14 mg·kg-1和0.12 mg·kg-1,降幅分别为22.22%和33.33%。
叶面喷施苹果酸不仅影响各器官中的Cd含量,也影响Cd在相邻器官间的转运效率。喷施苹果酸显著抑制了Cd从穗颈向籽粒、旗叶向穗节的转移。与对照相比,苹果酸处理下Cd的TF籽粒/穗颈分别下降了24.37% 和23.16%,TF穗节/旗叶分别下降了8.99% 和28.24%。
2.2 叶面喷施苹果酸对水稻各器官营养元素含量的影响Ca、Fe、Mn和Zn等营养元素的含量在水稻不同品种及不同器官间差异非常明显。Ca和Mn含量在不同器官中的高低次序为:旗叶最高,穗节和穗颈次之,籽粒最低(图 2A、图 2C);Fe含量的分布特征表现为旗叶最高,籽粒和穗颈次之,穗节最低(图 2B);Zn含量的分布特征表现为穗节最高,穗颈次之,籽粒和旗叶最低(图 2D)。营养元素在不同品种中分布也不同,X24和F36的Ca含量明显高于705和TGW,TGW的Mn含量明显低于其他3个品种,X24和705穗节中的Zn含量明显高于F36和TGW。

|
图 2 水稻开花期叶面喷施苹果酸对水稻各品种各器官中Ca、Fe、Mn、Zn含量及其平均含量增减幅度的影响 Figure 2 Effects of foliar application with malic acid at anthesis stage on the content of Ca, Fe, Mn and Zn in different organs, as well as increase percentage based on average value of 4 varieties |
喷施苹果酸后,4个品种各器官中的Mn、Fe和Zn平均含量均显著下降,但Ca的平均含量显著上升(图 2E),喷施2次和3次苹果酸使籽粒中的Ca含量分别增加16.02% 和26.66%,Fe含量分别减少30.54% 和43.14%,Mn含量分别减少23.07% 和28.55%,Zn含量分别减少11.31%和19.57%;穗颈中的Ca含量分别增加19.60% 和47.86%,Fe含量分别减少35.24% 和47.52%,Mn含量分别减少12.85% 和37.99%,Zn含量分别减少30.78%和50.28%;穗节中Ca含量分别增加8.67% 和31.25%,Fe含量分别减少22.27% 和41.57%,Mn含量分别减少15.78% 和31.67%,Zn含量分别减少17.64%和37.34%,旗叶中Ca含量分别增加4.41% 和36.26%,Fe含量分别减少57.37% 和81.59%,Mn含量分别减少24.05% 和29.13%,Zn含量分别减少18.09%和22.65%。
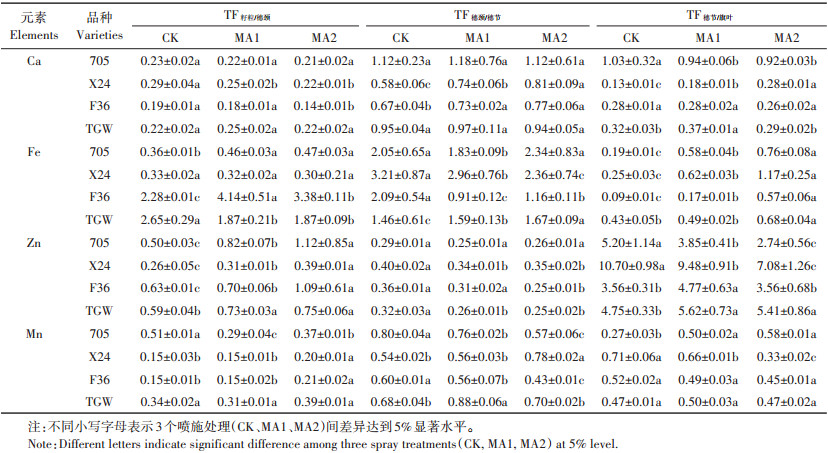
2.3 叶面喷施苹果酸对水稻各器官营养元素转移系数的影响叶面喷施苹果酸不仅影响各器官中必需营养元素的含量,也影响营养元素在相邻器官间的转运效率。苹果酸能促进Ca从穗节向穗颈中转移,而且各个品种之间也有差异,其中X24和F36的TF穗颈/穗节与CK差异达到显著水平;苹果酸也显著促进了Fe从旗叶向穗节的转移。与此同时,喷施苹果酸抑制了Zn从穗节向穗颈的转移,但促进了Zn从穗颈向籽粒中转移,705中Zn的TF籽粒/穗颈显著提高(表 2)。
|
|
表 2 苹果酸对水稻器官间营养元素转移系数的影响 Table 2 Effects of malic acid on the transfer factors of nutrient elements between rice organs |
稻米中的谷氨酸含量(18.37~24.34 mg·kg-1)显著高于其他氨基酸,约是其他氨基酸的2倍以上(图 3A)。在Cd污染土壤中进行栽培时,705和TGW籽粒中的谷氨酸含量明显低于X24和F36。亮氨酸、精氨酸、缬氨酸、甘氨酸、天冬氨酸、苯丙氨酸、赖氨酸、丝氨酸、胱氨酸、丙氨酸的含量在品种间的差异比较小,大多在3.49~8.52 mg·kg-1的范围之内。异亮氨酸、酪氨酸和组氨酸的含量相对较低,但它们在F36中的含量显著高于其他3个品种。喷施2次苹果酸对稻米中的氨基酸含量没有显著影响,喷施3次苹果酸显著提高了甘氨酸和缬氨酸的含量,与CK相比,甘氨酸和缬氨酸的含量分别增加了46.45% 和34.56%(图 3B)。喷施3次苹果酸对谷氨酸、亮氨酸和胱氨酸的含量也有明显的提高,但未达到5%的显著水平。

|
*表示处理间差异达到5%显著水平 * indicates significant difference between different treatments at 5% level 图 3 水稻开花期叶面喷施3次苹果酸对4个水稻品种籽粒氨基酸含量及其平均含量的影响 Figure 3 Effects of 3 times foliar application with malic acid at anthesis stage on the amino acid content and average content of 4 varieties |
苹果酸能通过呼吸作用对植物体内许多代谢过程产生调控作用。最近的研究表明,苹果酸具有信号传导功能,在提高植物抗逆能力方面发挥着重要作用[13, 32-33]。虽然水稻叶片中含有大量的苹果酸,但水稻开花期喷施5 mmol·L-1的苹果酸,依然能够对水稻灌浆过程中的Cd和营养元素的转运产生显著的影响[19]。近年来,叶面喷施肥料已经成为了水稻降Cd的重要措施。有研究表明,通过叶面喷施Zn和Mn等必需元素[34],能够提高水稻抗逆性,抑制Cd从根系向籽粒的转运;喷施黄腐酸、胡敏酸钾、有机硅等能够降低籽粒中的Cd含量[35-36]。本研究结果显示,叶面喷施苹果酸也能够降低稻米Cd含量,并且喷施两次5 mmol·L-1苹果酸的效果更好。
水稻籽粒中的Cd含量受基因型和环境的共同影响,品种类型、生长季节、海拔高度、播种方式、灌水方式等都会显著影响稻米中的Cd含量[7, 30, 37-38]。本研究选用的4个早稻品种Cd积累特性不同,在籽粒和穗颈中Cd含量由多到少依次为:705>X24>TGW>F36。对照处理下,稻米中的Cd含量(0.22~0.29 mg·kg-1)均大于《食品安全国家标准食品中污染物限量》(GB 2762—2017)规定的大米Cd含量上限0.2 mg·kg-1,各器官中的Cd含量也有明显的差异,其中穗节中的Cd含量最高,品种间的差异也最大,变幅为0.55~2.67 mg·kg-1。经过穗颈和穗轴的拦截,籽粒中的Cd含量在品种间的差异明显变小,喷施苹果酸后,4个品种籽粒中的Cd含量均下降至0.2 mg·kg-1以下,且降Cd幅度分别达到了46.15%和65.38%;与此同时,穗节中的Cd含量以及Cd在器官间的转移系数TF穗节/旗叶和TF籽粒/穗颈也大幅度下降。这可能是叶面喷施的苹果酸一部分与叶片中的Cd形成了MA-Cd螯合物,阻止了灌浆过程中叶片中的Cd向穗节的转运;另一部分可能转运到穗节和穗颈中,在这些器官中与Cd形成MA-Cd螯合物,抑制了Cd从穗颈向籽粒的转运,致使稻米中Cd的含量大幅度下降[39-40]。由此可见,喷施苹果酸能够有效抑制水稻灌浆期间Cd从旗叶向稻米中的转运,使Cd污染农田中的稻米Cd超标风险显著下降,从而实现污染农田安全利用的目标;但在营养生长阶段喷施苹果酸,是否能够抑制污染土壤中的Cd进入根系及其向营养器官转运,尚需进一步的研究。
非选择性阳离子通道(NSCCs)是一种普遍存在于生物膜上的离子通道,它们能够直接参与植物体内的许多生理过程,如摄取营养、维持压力、传导信号等[41-42]。水稻体内的NSCCs能优先转运必需元素,提高必需元素Ca、Mn、Zn的含量或者抑制NSCCs的活性,都能有效削弱Cd对NSCCs的竞争性结合,显著抑制水稻植株对Cd的吸收和转运[35, 43-44]。离子通道的开放与关闭状态常受许多因素的影响,一般将离子通道分为电压门控型、配体门控型和机械门控型三大类[45-46],但电压、配体和机械刺激间有复杂的交互作用[47]。本研究发现,叶面喷施苹果酸,能同时降低水稻籽粒、穗颈和穗节中的Cd含量以及Fe、Mn、Zn的含量,却显著提高了这些器官中的Ca含量(图 2E)。说明苹果酸对离子通道的开放程度和选择透性产生了调控作用,使得Zn、Fe、Mn和Cd的转运受到抑制,导致它们在各器官中的含量下降;或者是苹果酸对锌铁蛋白(ZIP)的离子转运功能产生了抑制作用[44, 48],致使穗节等器官中的Zn、Fe和Cd同步下降(图 2B、图 2D)。苹果酸在降低Cd、Fe、Mn和Zn离子转运速率的同时,也提高了Ca2+从稻穗各部位向籽粒中的转运。大量的研究证明,Ca2+是生物体内最重要的信号因子之一[41],Ca2+在动物、植物和微生物抵制不良环境过程中发挥了重要的作用[49-51]。有些离子通道对重金属的敏感性既与离子通道的氨基酸残基构成有关,也与细胞中的Ca浓度有关[52-53]。苹果酸有可能通过增加水稻穗部器官中Ca2+的浓度提高离子通道对有害重金属Cd的识别和拦截能力。
氨基酸是合成蛋白质最重要的前体物质和组分[54],与酶活性、基因表达和离子转运的调控有关,在提高植物抗逆能力方面发挥着关键作用[55]。Cd污染不仅会降低水稻各个器官必需元素的含量,而且会导致籽粒中氨基酸含量的下降[22]。本研究发现,稻米中的谷氨酸含量远高于其他氨基酸,在品种间有显著的差异,叶面喷施3次苹果酸能够显著提高甘氨酸和缬氨酸的含量,对谷氨酸、亮氨酸和胱氨酸的合成也有明显的促进作用。这可能是因为叶面喷施苹果酸后,稻米的Cd含量大幅度下降,Cd对氨基酸合成的抑制效应被消除;也可能是苹果酸通过促进TCA循环,提高α-酮戊二酸向谷氨酸的转化效率,进而促进了其他氨基酸的合成[56-57]。
4 结论(1)水稻开花期喷施2~3次苹果酸能显著降低成熟期籽粒、穗颈、旗叶和穗节中的Cd含量,对4个水稻品种全部有效。喷施苹果酸后经过穗颈和穗轴的拦截,4个品种籽粒中的平均Cd含量从0.26 mg·kg-1分别下降到0.14 mg·kg-1和0.09 mg·kg-1,降幅分别为46.15%和65.38%。
(2)叶面喷施苹果酸对水稻各器官中Ca、Fe、Mn、Zn的含量及其在相邻器官间的转移系数产生了显著影响。喷施苹果酸后,各器官中的Mn、Fe和Zn含量均显著下降,Ca的含量显著上升。
(3)喷施3次苹果酸后,稻米中的甘氨酸和缬氨酸含量分别增加了46.45%和34.56%;对谷氨酸、亮氨酸和胱氨酸的合成也有明显的促进作用。
| [1] |
卢夏茹. 普通野生稻种子贮藏蛋白质多态性研究[D]. 海口: 海南大学, 2015. LU Xia-ru. Study on polymorphism of common wild rice seed storage protein[D]. Haikou: Hainan Unitersity, 2015. |
| [2] |
Kerkhove E V, Valérie P, Swennen Q. Cadmium and transport of ions and substances across cell membranes and epithelia[J]. Biogeotechnology of Metals, 2010, 23(5): 823-855. |
| [3] |
Bermudez G M A, Jasan R, Rita Plá, et al. Heavy metals and trace elements in atmospheric fall-out: Their relationship with topsoil and wheat element composition[J]. Journal of Hazardous Materials, 2012, 213/214: 447-456. DOI:10.1016/j.jhazmat.2012.02.023 |
| [4] |
He S, He Z, Yang X, et al. Soil biogeochemistry, plant physiology, and phytoremediation of cadmium contaminated soils[J]. Advances in Agronomy, 2015, 134: 135-225. |
| [5] |
Song W E, Shi B, et al. Variation of Cd concentration in various rice cultivars and derivation of cadmium toxicity thresholds for paddy soil by species-sensitivity distribution[J]. Journal of Integrative Agriculture, 2015(9): 1845-1854. |
| [6] |
刘洋. 水稻非选择性阳离子通道离子转运效率的遗传多样性分析[D]. 天津: 天津农学院, 2017. LIU Yang. Genetic diversity analysis of ion transport efficiency in rice non-selective cation channels[D]. Tianjin: Tianjin Agricultural College, 2017. |
| [7] |
黄永春, 张长波, 任兴华, 等. 土壤和茎基部Cd含量对稻米Cd污染风险的影响[J]. 农业环境科学学报, 2020, 39(5): 989-999. HUANG Yong-chun, ZHANG Chang-bo, REN Xing-hua, et al. Effects of cadmium content in soil and stem base on the risk of cadmium contamination in rice[J]. Journal of Agro-Environment Science, 2020, 39(5): 989-999. |
| [8] |
吴清平, 周小燕. L-苹果酸研究进展[J]. 微生物学通报, 1990, 17(1): 30-33. WU Qing-ping, ZHOU Xiao-yan. Research progress of physiological functions of L-malate[J]. Microbiology China, 1990, 17(1): 30-33. |
| [9] |
Chen Q Q, Wang B P, Ding H Y, et al. Review: The role of NADP-malic enzyme in plants under stress[J]. Plant Science, 2019, 281: 206-212. DOI:10.1016/j.plantsci.2019.01.010 |
| [10] |
李秀峰, 张欣欣, 柳参奎, 等. 水稻苹果酸酶基因在逆境下的表达特性研究[J]. 基因组学与应学, 2012, 31(4): 327-332. LI Xiufeng, ZHANG Xin-xin, LIU Can-kui, et al. Study on the expression characteristics of malatase gene in rice under stress[J]. Genomics and Applied Biology, 2012, 31(4): 327-332. |
| [11] |
Tao P, Guo W P, Li B Y, et al. Genome-wide identification, classification, and analysis of NADP-ME family members from 12 crucifer species[J]. Molecular Genetics and Genomics, 2016, 291(3): 1167-1180. DOI:10.1007/s00438-016-1174-3 |
| [12] |
Sun X, Han G L, Meng Z, et al. Roles of malic enzymes in plant development and stress responses[J]. Plant Signaling and Behavior, 2019, 14(10): 1-8. |
| [13] |
Bela K, Horváth E, Gallé Á, et al. Plant glutathione peroxidases: Emerging role of the antioxidant enzymes in plant development and stress responses[J]. Journal of Plant Physiology, 2015, 176: 192-201. DOI:10.1016/j.jplph.2014.12.014 |
| [14] |
Wang S T, Dong Q, Wang Z L. Differential effects of citric acid on cadmium uptake and accumulation between tall fescue and Kentucky bluegrass[J]. Ecotoxicology and Environmental Safety, 2017, 145: 200-206. DOI:10.1016/j.ecoenv.2017.07.034 |
| [15] |
Hawrylak-Nowak B, Dresler S, Matraszek R. Exogenous malic and acetic acids reduce cadmium phytotoxicity and enhance cadmium accumulation in roots of sunflower plants[J]. Plant Physiology and Biochemistry, 2015, 94: 225-234. DOI:10.1016/j.plaphy.2015.06.012 |
| [16] |
Mnasri M, Ghabriche R, Fourati E, et al. Cd and Ni transport and accumulation in the halophyte Sesuvium portulacastrum: Implication of organic acids in these processes[J]. Frontiers in Plant Science, 2015, 6: 156-160. |
| [17] |
Eniko T, Viktor G M, Anita V, et al. Effect of lead nickel and vanadium contamination on organic acid transport in xylem sap of cucumber[J]. Journal of Inorganic Biochemistry, 1999, 75(3): 219-223. DOI:10.1016/S0162-0134(99)00091-4 |
| [18] |
Sebastian A, Prasad M N V. Exogenous citrate and malate alleviate cadmium stress in Oryza sativa L.: Probing role of cadmium localization and iron nutrition[J]. Ecotoxicology and Environmental Safety, 2018, 166: 215-222. DOI:10.1016/j.ecoenv.2018.09.084 |
| [19] |
张烁, 陆仲烟, 唐琦, 等. 水稻叶面调理剂的降Cd效果及其对营养元素转运的影响[J]. 农业环境科学学报, 2018, 37(11): 2507-2513. ZHANG Shuo, LU Zhong-yan, TANG Qi, et al. Effect of leaf conditioner on reducing Cd and its effect on nutrient transport in rice[J]. Journal of Agro-Environment Science, 2018, 37(11): 2507-2513. |
| [20] |
Li T, Di Z, Islam E, et al. Rhizosphere characteristics of zinc hyperaccumulator Sedum alfredii involved in zinc accumulation[J]. Journal of Hazardous Materials, 2011, 185(2): 818-823. |
| [21] |
Wei S, Guo B, Feng L, et al. Cadmium distribution and characteristics of cadmium-binding proteins in rice(Oryza sativa L.)Kernel[J]. Food Science and Technology Research, 2017, 23(5): 661-668. DOI:10.3136/fstr.23.661 |
| [22] |
Yuan K, Wang C R, Zhang C B, et al. Rice grains alleviate cadmium toxicity by expending glutamate and increasing manganese in the cadmium contaminated farmland[J]. Environmental Pollution, 2020, 262: 1-10. |
| [23] |
Zhao Y L, Zhang C B, Wang C R, et al. Increasing phosphate inhibits cadmium uptake in plants and promotes synthesis of amino acids in grains of rice[J]. Environmental Pollution, 2020, 257: 113496. DOI:10.1016/j.envpol.2019.113496 |
| [24] |
Chen J, Shafi M, Wang Y, et al. Organic acid compounds in root exudation of moso bamboo(Phyllostachys pubescens)and its bioactivity as affected by heavy metals[J]. Environmental Science and Pollution Research, 2016, 23(20): 20977-20984. DOI:10.1007/s11356-016-7323-8 |
| [25] |
Vecerova K, Vecera Z, Docekal B, et al. Changes of primary and secondary metabolites in barley plant exposed to CdO nanoparticles[J]. Environmental Pollution, 2016, 218: 207-218. DOI:10.1016/j.envpol.2016.05.013 |
| [26] |
Montiel R, Madejón E, Madejón P. Effect of heavy metals and organic matter on root exudates(low molecular weight organic acids)of herbaceous species: An assessment in sand and soil conditions under different levels of contamination[J]. Environmental Pollution, 2016, 216: 273-281. DOI:10.1016/j.envpol.2016.05.080 |
| [27] |
居学海, 张长波, 宋正国, 等. 水稻籽粒发育过程中各器官Cd积累量的变化及其与基因型和土壤Cd水平的关系[J]. 植物生理学报, 2014, 50(5): 634-640. JU Xue-hai, ZHANG Chang-bo, SONG Zheng-guo, et al. Changes in cadmium accumulation in rice organs during grain development and their relationship with genotype and cadmium levels in soil[J]. Plant Physiology Journal, 2014, 50(5): 634-640. |
| [28] |
文志琦, 赵艳玲, 崔冠男, 等. 水稻营养器官Cd积累特性对稻米Cd含量的影响[J]. 植物生理学报, 2015, 51(8): 1280-1286. WEN Zhi-qi, ZHAO Yan-ling, CUI Guan-nan, et al. Effects of accumulation characteristics of cadmium in rice vegetative organs on cadmium content in rice[J]. Plant Physiology Journal, 2015, 51(8): 1280-1286. |
| [29] |
王龙, 高子平, 李文华, 等. 水稻幼苗Cd吸收动力学特性的遗传多样性分析[J]. 植物生理学报, 2016, 52(1): 125-133. WANG Long, GAO Zi-ping, LI Wen-hua, et al. Genetic diversity of cadmium absorption kinetic characteristics in rice(Oryza sativa) seedlings[J]. Plant Physiology Journal, 2016, 52(1): 125-133. |
| [30] |
Liu Y, Zhang C B, Zhao Y L, et al. Effects of growing seasons and genotypes on the accumulation of cadmium and mineral nutrients in rice grown in cadmium contaminated soil[J]. Science of the Total Environment, 2017, 579: 1282-1288. DOI:10.1016/j.scitotenv.2016.11.115 |
| [31] |
Shi G L, Zhu S, Bai S N, et al. The transportation and accumulation of arsenic, cadmium, and phosphorus in 12 wheat cultivars and their relationships with each other[J]. Journal of Hazardous Materials, 2015, 299: 94-102. DOI:10.1016/j.jhazmat.2015.06.009 |
| [32] |
Finkemeier I, Konig A C, Heard W, et al. Transcriptomic analysis of the role of carboxylic acids in metabolite signaling in Arabidopsis leaves[J]. Plant Physiology Journal, 2013, 162(1): 239-253. DOI:10.1104/pp.113.214114 |
| [33] |
Zhao Y, Luo L, Xu J. Malate transported from chloroplast to mitochondrion triggers production of ROS and PCD in Arabidopsis thaliana[J]. Circulation Research, 2018, 28(4): 448-461. |
| [34] |
韩潇潇, 任兴华, 王培培, 等. 叶面喷施锌离子对水稻各器官Cd积累特性的影响[J]. 农业环境科学学报, 2019, 38(8): 1809-1817. HAN Xiao-xiao, REN Xing-hua, WANG Pei-pei, et al. Effects of foliar application with zinc on the characteristics of cadmium accumulation in organs of rice plants[J]. Journal of Agro-Environment Science, 2019, 38(8): 1809-1817. |
| [35] |
Wang S, Wang F, Gao S, et al. Heavy metal accumulation in different rice cultivars as influenced by foliar application of nano-silicon[J]. Water Air and Soil Pollution, 2016, 227(7): 1-13. DOI:10.1007/s11270-016-2928-6 |
| [36] |
Chen R, Zhang C B, Liu Z Q, et al. Foliar application with nano-silicon reduced cadmium accumulation in grains by inhibiting cadmium translocation in rice plants[J]. Environmental Science and Pollution Research, 2018, 25(3): 2361-2368. DOI:10.1007/s11356-017-0681-z |
| [37] |
Deng X, Yang Y, Zeng H, et al. Variations in iron plaque, root morphology and metal bioavailability response to seedling establishment methods and their impacts on Cd and Pb accumulation and translocation in rice(Oryza sativa L.)[J]. Journal of Hazardous Materials, 2020, 384: 121343. DOI:10.1016/j.jhazmat.2019.121343 |
| [38] |
Spanu A, Valente M, Langasco I, et al. Sprinkler irrigation is effective in reducing cadmium concentration in rice(Oryza sativa L.)grain: A new twist on an old tale?[J]. Science of the Total Environment, 2018, 628: 150-160. |
| [39] |
Han Y, Huang S, Yuan H, et al. Organic acids on the growth, anatomical structure, biochemical parameters and heavy metal accumulation of Iris lactea var. Chinensis seedling growing in Pb mine tailings[J]. Ecotoxicology, 2013, 22(6): 1033-1042. DOI:10.1007/s10646-013-1089-2 |
| [40] |
Chigbo C, Batty L. Chelate-assisted phytoremediation of Cu-pyrenecontaminated soil using Z. mays[J]. Water Air and Soil Pollution, 2015, 226(3): 74-83. DOI:10.1007/s11270-014-2277-2 |
| [41] |
刘仲齐, 张长波. 重金属调控非选择性阳离子通道生理功能的研究进展[J]. 农业资源与环境学报, 2017, 34(1): 1-5. LIU Zhongqi, ZHANG Chang-bo. Research progress of heavy metals in regulating the physiological functions of non-selective cation channels[J]. Journal of Agricultural Resources and Environment, 2017, 34(1): 1-5. |
| [42] |
Pinto E, Ferreira I. Cation transporters/channels in plants: Tools for nutrient biofortification[J]. Journal of Plant Physiology, 2015, 179: 64-82. DOI:10.1016/j.jplph.2015.02.010 |
| [43] |
Han X X, Zhang C B, Wang C R, et al. Gadolinium inhibits cadmium transport by blocking non-selective cation channels in rice seedlings[J]. Ecotoxicology and Environmental Safety, 2019, 179(15): 160-166. |
| [44] |
张参俊, 张长波, 王景安, 等. 非选择性阳离子通道对水稻幼苗Cd吸收转运特性的影响[J]. 农业环境科学学报, 2015, 34(6): 1028-1033. ZHANG Shen-jun, ZHANG Chang-bo, WANG Jing-an, et al. Effects of nonselective cation channels on Cd accumulation and transfer characteristics in rice seedling[J]. Journal of Agro-Environmental Science, 2015, 34(6): 1028-1033. |
| [45] |
Demidchik V, Davenport R J, Tester M. Nonselective cation channels in plants[J]. Annual Review of Plant Biology, 2002, 53: 67-107. DOI:10.1146/annurev.arplant.53.091901.161540 |
| [46] |
刘胜浩, 刘晨临, 黄晓航, 等. 植物细胞的非选择性阳离子通道[J]. 植物生理学通讯, 2006, 42(3): 523-528. LIU Sheng-hao, LIU Chen-lin, HUANG Xiao-hang, et al. Nonselective cation channels in plant cells[J]. Plant Physiology Communications, 2006, 42(3): 523-528. |
| [47] |
刘仲齐, 张长波, 黄永春. 水稻各器官Cd阻控功能的研究进展[J]. 农业环境科学学报, 2019, 38(4): 721-727. LIU Zhong-qi, ZHANG Chang-bo, HUANG Yong-chun. Research advance on the functions of rice organs in cadmium inhibition: A review[J]. Journal of Agro-Environment Science, 2019, 38(4): 721-727. |
| [48] |
尹洁, 赵艳玲, 高子平, 等. 锌对粳稻幼苗镉吸收转运特性的影响[J]. 农业环境科学学报, 2016, 35(5): 834-841. YIN Jie, ZHAO Yan-ling, GAO Zi-ping, et al. Effects of zinc supply on absorption and translocation of cadmium in rice seedlings[J]. Journal of Agro-Environment Science, 2016, 35(5): 834-841. |
| [49] |
Bouron A, Kiselyov K, Oberwinkler J. Permeation, regulation and control of expression of TRP channels by trace metal ions[J]. European Journal of Applied Physiology, 2015, 467: 1143-1164. |
| [50] |
Hepler P K. Calcium: A central regulator of plant growth and development[J]. Plant Cell, 2005, 17: 2142-2155. DOI:10.1105/tpc.105.032508 |
| [51] |
Jones B L, Smith S M. Calcium-sensing receptor: A key target for extracellular calcium signaling in neurons[J]. Frontiers in Physiology, 2016, 7: 116-123. |
| [52] |
Inoue R, Okada T, Onoue H, et al. The transient receptor potential protein homologue TRP6 is the essential component of vascular α1-adrenoceptor-activated Ca2+-permeable cation channel[J]. Circulation Research, 2001, 88(3): 325-332. DOI:10.1161/01.RES.88.3.325 |
| [53] |
Zhou Y, Xia X M, Lingle C J. Cadmium-cysteine coordination in the B K inner pore region and its structural and functional implications[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(16): 5237-5242. DOI:10.1073/pnas.1500953112 |
| [54] |
王莹, 史振声, 王志斌, 等. 植物对氨基酸的吸收利用及氨基酸在农业中的应用[J]. 中国土壤与肥料, 2008(1): 6-11. WANG Ying, SHI Zhen-sheng, WANG Zhi-bin, et al. Absorption and utilization of amino acids in plants and their application in agriculture[J]. Soil and Fertilizer Sciences in China, 2008(1): 6-11. |
| [55] |
Cuin T A, Shabala S. Amino acids regulate salinity-induced potassium efflux in barley root epidermis[J]. Planta, 2007, 225(3): 753-761. DOI:10.1007/s00425-006-0386-x |
| [56] |
Forde B G, Lea P J. Glutamate in plants: Metabolism, regulation, and signaling[J]. Journal of Experimental Botany, 2007, 58(9): 2339-2358. DOI:10.1093/jxb/erm121 |
| [57] |
Galili G, Amir R, Fernie A R. The regulation of essential amino acid synthesis and accumulation in plants[J]. Annual Review of Plant Biology, 2016, 67(1): 153-178. DOI:10.1146/annurev-arplant-043015-112213 |